
Diversidad de Rotíferos Monogonta del Litoral Fluvial Argentino
Susana JOSÉ de PAGGI1
1Instituto Nacional de Limnología. CONICET-UNL. Santo Tomé, Santa Fe.
Abstract: Current knowledge on rotifer fauna of the Argentinean Fluvial Litoral is commented. This study mainly deals with Paraná River fauna, because the knowledge about Uruguay River is scarce. This contribution is based upon information published in taxonomic and faunistic papers and the author's unpublished data. Rotifer fauna of the Paraná River includes nearly 250 morphospecies, belonging to 21 families and 44 genera. The most diverse genera are Lecane, Trichocerca and Brachionus . In the flood-plain shallow lakes above 225 species were found, while 113 species were recorded in the main channel. The local diversity can be variable, depending on location of the lakes in the plain, connection to the river, environmental complexity, biotic interactions and methodological aspects.
Key words: Rotífera, Argentinean Mesopotamia, Taxonomy
Palabras clave: Rotifera, Mesopotamia Argentina, Taxonomía
Los rotíferos son organismos microscópicos, acuáticos y semiacuáticos, más conocidos en la limnologia por ser componentes del plancton ( microplancton ) aunque están muy bien representados en las comunidades litorales y también forman parte del zoomicrobentos. La mayoría es de vida libre, hay pocos parásitos, generalmente son solitarios, pero hay especies que forman colonias de variable tamaño.
Son organismos muy diversos en formas que se caracterizan por dos elementos particulares como son la corona de cilios de la cabeza y el trophi o estructura masticatoria. Constituyen comparativamente un pequeño filum pero son muy importantes en los ambientes continentales a causa de su tasa reproductora, su habilidad para ocupar rápidamente los nichos vacantes, constituyendo más del 30% de la biomasa planctonica, son recicladores eficientes de la materia orgánica y responden rápidamente a los cambios ambientales (Gannon et al., 1978, Nogrady et al., 1993) .
La gran mayoría ocupa aguas continentales ya que son comparativamente muy pocas las especies marinas. Colonizan ambientes con distinto grado de salinidad, pH y temperatura, tolerando muchos de ellos concentraciones muy bajas de oxígeno.
En general se reproducen por partenogénesis, producen machos por muy breves períodos, en condiciones desfavorables dan lugar a huevos de resistencia; hay casos comprobados de huevos que han eclosionado 20 años después.
Actualmente se considera que existen unas 2000 especies. Los rotíferos dulceacuícolas se reúnen en dos grandes Sub-clases Monogonta y Bdelloidea. La Sub- clase Monogonta es la más importante en los ambientes acuáticos continentales, reúne 95 géneros y 1600 especies (Nogrady et al., 1993, Segers 2002).
Estado actual de la Taxonomía del grupo
La taxonomía de los rotíferos se basa en el análisis de caracteres morfológicos externos e internos ( trophi ), evaluados a distinto nivel entre los que se incluyen análisis biométricos y la microscopia electrónica de barrido. Desde algunos años estos análisis han comenzado lentamente a completarse con estudios de cariotipos, empleo de electroforesis para el análisis de las proteínas, y secuenciación de ADN (Gómez 1998, Welch & Meselson 1998, entre otros).
Sin embargo, a pesar de estas modernas herramientas, que lejos están de constituir lo usual, la taxonomía de los rotíferos presenta diversos problemas largamente analizados (Koste & Shiel 1989, Ruttner-Kolisko 1989, Nogrady et al., 1993) las causas de los mismos pueden agruparse en: partenogénesis facultativa u obligada, lo cual implica transferencia de genotipos de padres a hijos y por lo tanto transferencia de mutaciones que cuando no son letales se multiplican rápidamente en la población y generan un alto grado de variabilidad; corto tiempo de generaciones, se estima unas 100 por año bajo condiciones favorables, lo que contribuye a multiplicar cualquier cambio genético; insuficiencia de caracteres morfológicos útiles, lo cual junto con la variabilidad es la dificultad más seria en la taxonomía del grupo.
La alta variabilidad de algunos caracteres y variaciones fenotípicas (inducidas por la dieta, por la depredación y eclosión de huevos de resistencia) constituyen verdaderos problemas en el contexto de una taxonomía basada en la morfología externa. Otra cuestión no menos importante es la existencia demostrada en la última década de sibling especies entre los rotíferos (Fu et al.,1991, Gómez & Serra 1995; Ciros-Perez et al., 2001).
Estado actual del conocimiento de la Rotiferofauna del Litoral
La información disponible sobre los rotíferos de la región del litoral fluvial argentino proviene de estudios faunísticos realizados principalmente en determinados sectores de la cuenca del Paraná (diversas publicaciones en José de Paggi 1990; José de Paggi, 1996, 2001, 2002). En varios trabajos de ecología del zooplancton se incluyen también inventarios de especies, que dan una estimación general de la riqueza del filum (José de Paggi, 1980, 1984, Bonetto & Corrales de Jacobo 1985, Frutos 1996, 1998 entre otros).
Sin duda la riqueza potencial de los rotíferos en la cuenca del Paraná puede ser más alta que la actualmente conocida. La complejidad del sistema a escala temporal y espacial dada por la extensión de su planicie y el régimen de pulsos (Neiff 1990) favorece una alta diversidad biológica. Por otro lado hay secciones de la cuenca muy poco exploradas como los del Paraná inferior y su delta (Modenutti & Claps 1988).
Es muy poca la información publicada sobre los rotíferos de la otra gran cuenca hidrográfica del litoral, la del río Uruguay (Marinone 1994), un muestreo realizado en algunos de los brazos de la represa de Salto Grande puso en evidencia la existencia en Argentina de un rotífero invasor, Kellicottia bostoniensis (José de Paggi 2002). Son escasos los estudios sobre la fauna de rotíferos de ambientes acuáticos lénticos y lóticos del interior de las provincias de Entre Ríos y Misiones. La información disponible sobre algunos ambientes de la provincia de Corrientes dan cuenta de la alta riqueza del grupo (José de Paggi 1996).
Finalmente debe señalarse también que una parte importante de la información publicada con que se cuenta está sesgada por cuestiones estrictamente taxonómicas y metodológicas. Acerca de muchas especies no se tiene certeza en cuanto a una determinacion taxonómica segura. Por ejemplo para el género Trichocerca la información disponible señala la existencia de 27 especies para la cuenca del Paraná, sin embargo sólo la mitad de estas fueron clasificadas en el marco de trabajos de tipo taxonómico y sólo para cuatro de ellas se cuenta con observaciones particulares sobre su morfología, dimensiones y registros gráficos que hacen posible que cualquier investigador sepa a que especie se refieren. El resto corresponde a especies incluídas en inventarios de trabajos ecológicos. Tratándose de un género en que es imprescindible para una buena identificación la observación de los caracteres de las piezas del trophi , no hay certeza acerca de las determinaciones realizadas en tales trabajos.
Diversidad gamma
El total de rotíferos planctónicos y litorales de la cuenca del Paraná entendido como diversidad regional o diversidad gamma (Ricklefs 1998) es aproximadamente de unas 250 especies pertenecientes a 21 familias y 44 géneros (Tabla 1). Representan el 72 y 41% de los conocidos respectivamente para el grupo (Segers 2002). Esta información proviene de un total de estudios publicados e inéditos en el que se incluyen unas 80 lagunas distribuidas desde la confluencia del Paraná con el río Paraguay, hasta su desembocadura y unos 15 ambientes lóticos (curso principal del río Paraná, muestreado por períodos consecutivos de casi siete años y cauces secundarios y tributarios). La riqueza de rotíferos de la planicie, pese a contar con una comparativamente menor intensidad de muestreos, es mayor que la registrada en los ambientes lóticos. Mientras que en los ambientes de la planicie se han registrado unas 225 especies, en el curso principal del Paraná se registraron 113 especies. También es mayor el número de familias y géneros que se ha observado en los ambientes de la planicie.
Las familias mas ricamente representadas son Lecanidae Remane, Trichocercidae Harring y Brachionidae Ehrenberg. El género Lecane reúne poco menos de 50 especies, lo cual constituye casi un tercio del total de las especies del género. Trichocerca estaría representada por casi 30 especies de las 65 que posee el género (Figs. 1 y 2). La riqueza del género Brachionus , unas 20 especies, es muy alta comparándola con otras cuencas hidrográficas de Argentina (José de Paggi 1990).
Las especies de estos géneros mencionados se encuentran formando parte del plancton de las lagunas someras, y aún de los ríos, pero la mayor diversidad se registra en la zona litoral vegetada de las lagunas. Trichocerca juntamente con el género Lecane constituyen los de mayor diversidad en los ambientes de ríos con planicie aluvial (Segers et al., 1993)
La riqueza de los géneros Lecane y Brachionus se relacionaría por un lado a los requerimientos ecológicos de las especies y por otro lado a la ubicación biogeográfica de la cuenca del Paraná en la subregión Guayano brasileña, donde ambos géneros detectan un gran riqueza (Koste & José de Paggi 1982).
Otras familias como Testudinellidae Harring y Lepadellidae Harring también se encuentran comparativamente bien representadas, a nivel de los géneros Testudinella y Lepadella .
A medida que se incrementan los estudios faunísticos se registran nuevos géneros para Argentina e incluso especies nuevas para la ciencia, como así también se amplía el conocimiento sobre la diversidad de géneros litorales como los pertenecientes a las familias Notommatidae Hudson & Gosse y Dicranophoridae Harring (José de Paggi 2001ab).
Diversidad alfa
El número de especies por hábitat acuático del sistema del Paraná o diversidad alfa puede ser muy variable, dependiendo de su ubicación en la llanura, el estado de conexión al río, desarrollo de su litoral vegetado, interacciones bióticas y cuestiones metodológicas como número de estaciones de muestreo, diámetro de malla de redes, etc. La complejidad estructural de las lagunas incrementada por su área litoral puede sustentar una muy rica diversidad de rotíferos. El siguiente ejemplo puede ser ilustrativo de ello, las lagunas La Sirena y El Tigre fueron muestreadas por coincidencia durante un mismo número de meses (14), la primera de 200 hectareas, ubicada en la estrecha planicie izquierda del Alto Paraná, provincia de Corrientes y la segunda de sólo 3 hectáreas ubicada próxima a Santa Fe. En la Sirena se consideró solamente una estación limnética y se registraron 55 especies de rotíferos (Corrales de Jacobo & Frutos 1985) mientras que en El Tigre se consideraron además de una estación limnética, 4 ubicadas en la región litoral. Se hallaron 111 especies (Jose de Paggi, 1993).
Las localidades estructuralmente más complejas, permiten la coexistencia de un mayor número de especies por segregación de hábitas (Menge & Sutherland, 1976). Dentro de una misma laguna número y composición de especies pueden diferir según se considere el área pelágica y litoral. Así en estudios realizados en 6 lagunas ubicadas a lo largo del Paraná en los primeros tramos aguas abajo de su confluencia con el Paraguay (José de Paggi, inédito), el total de especies dentro de cada área experimentó variaciones numéricas, la composición tambien fue diferente, lo cual queda expresado en una afinidad faunística no mayor del 45 % (tabla 2). Mientras que en las áreas pelágicas la rotiferofauna se caracteriza por dominancia de especies de géneros planctónicos como Keratella, Polyarthra, Filinia en las áreas litorales se registra un numero mayor de géneros y dominancia de especies de Lecane, Trichocerca, Lepadella. Por otro lado las lagunas con mayor cobertura de vegetación detectaron en este grupo una mayor riqueza general de especies.
La diversidad alfa muestra cambios en relación a la estacionalidad hidrológica del Paraná, el pulso de inundación que conecta río y planicie, favorece el intercambio y redistribución de especies entre los ambientes acuáticos. Así por ejemplo en un grupo de 11 lagunas en islas próximas a Santa Fe (José de Paggi, inédito) la diversidad alfa promedio fue de 11 (CV: 53) y de 15 (CV: 46) antes y después de un evento de inundación. En el 82% de las lagunas la afinidad faunística entre ellas antes y después de la inundación fue menor del 40% (coeficiente de Jaccard) señalando la intensidad del cambio en la composición de las especies.
La riqueza local o diversidad alfa para un determinado momento, puede variar mucho en los ambientes lénticos de la cuenca del Paraná. Desde muy pocas especies, 3 o 4 hasta un total de 51 y 56 especies como los hallados en las lagunas Junco y Concepción, respectivamente ubicadas próximas a la desembocadura del río Corrientes con el Paraná (José de Paggi 1996). Las lagunas del litoral fluvial probablemente se encuentren entre los ambientes acuáticos de mayor riqueza de especies de rotíferos de Argentina.
Algunas consideraciones
Un balance adecuado del grado de conocimiento actual de los rotíferos del litoral fluvial debe contemplar la premisa de que en las estimaciones de riqueza de cualquier grupo de plantas o animales debe tenerse muy en cuenta que éstas son altamente dependientes de la escala espacial de muestreo , intensidad del análisis y competencia taxonómica del investigador (Brown 1990; Kalff 2002).
No hay dudas de la importancia de la necesidad de continuar los estudios ajustándolos a la escala temporal y espacial de variación que presentan los grandes humedales del litoral. Pero la intensidad del análisis no debe referirse solamente al número adecuado de muestras sino también al nivel de resolución taxonómica que se necesita para la evaluación de la riqueza de especies. Si bien algunos autores muestran que en estudios ecológicos se pueden obtener resultados significativos a partir de una precisión taxonómica a nivel de familia y género (Clarke & Warwick 1994), los análisis realizados con rotíferos precisamente indican que solo la máxima resolución taxonómica, es decir a nivel de especies, puede contribuir a una evaluación integral de la biodiversidad (Nielsen et al., 1998).
Finalmente debe estimularse la formación de una buena competencia taxonómica. La sistemática es la base científica y de credibilidad para la obtención de buenos inventarios de biodiversidad (Wheeler 1995). Los tópicos de moda en investigación frecuentemente han desplazado los estudios taxonómicos como objeto de muchos proyectos científicos. Paradójicamente esto se ha dado en momentos en que este tipo de estudios son urgentes debido a que la destrucción de muchos hábitats naturales ha generado la llamada crisis de biodiversidad que implica que muchos organismos estén en peligro de extinción antes de haber sido clasificados (Quicke 1993;Prance 1995).
Bibliografía
Bonetto, A.A. & Corrales de Jacobo, M., 1985. Zooplancton del río Paraná medio: variaciones temporales y distribucionales en el área de confluencia con el río Paraguay. Ecosur 23/24: 1-23.
Brown, J.H., 1990. Species diversity, p 57-89. En: Myers, A. & Giller, P. (eds.) Analytical Biogeography.An integrate approach to the study of animal and plant distributions. Chapman & Hall.
Ciros-Perez, J., Gomez, A. & Serra, M., 2001. On the taxonomy of three sympatric sibling species of the Brachionus plicatilis (Rotifera) complex from Spain, with the description of B.ibericus n.sp. J. Plankton Res 23(12): 1311-1328.
Clarke, K. R. & Warwick, M., 1994. Changes in marine communities: an approach to statistical analysis and interpretation. Nat. Env. Res. Council. UK . 144 pp
Frutos, S.M., 1993 Zooplancton en cuerpos de agua isleños del Bajo Paraná. Ambiente subtropical 3: 87-121.
Fu,Y. Hirayama, K. & Natsukari, Y., 1991. Morphological differences between two types of the rotifer Brachionus plicatilis O.F. Muller. J.exp.mar.Biol.Ecol. 151: 29-41.
Gannon, J. E. & Stemberger, R.S., 1978. Zooplankton (especially crustaceans and rotifers) as indicators of water quality. Trans. am.Micros.Soc . 97: 16-35.
Gomez, A. & Serra, M., 1995. Behavioral reproductive isolation among sympatric strains of Brachionus plicatilis Muller, 1786: insights into the status taxonomic species. Hydrobiologia 313/314:: 111-119.
Gomez, A., 1998. Allozyme electrophoresis: its application to rotifers. Hydrobiologia 387/388: 385-393.
José de Paggi, S., 1980. Campaña limnológica Keratella I en el río Paraná medio. Zooplancton de ambientes loticos. Ecología 4: 69-75.
Koste, W. & José de Paggi, S., 1982. Rotifera of the Superorder Monogononta recorded from Neotropis. Gewäss.Abwäss. 68-69: 71-102.
José de Paggi, S.,1984. Estudios limnológicos en una sección transversal del tramo medio del río Paraná: distribución estacional del zooplancton. Revista Asociación Ciencias Naturales del Litoral 15(2): 135-155.
José de Paggi, S., 1990. Ecological and biogeographical remarks on the rotifer fauna of Argentina. Rev. Hydrobiol. trop. 23 (4): 297-311.
José de Paggi, S. ,1993.Composition and seasonality of planktonic rotifers in limnetic and littoral regions of a floodplain lake (Parana river system). Rev. Hydrobiol. trop . 26: 53-63.
José de Paggi, S., 1996. Rotifera (Monogononta) diversity in subtropical waters of Argentina. Annlales Limnología 32 (4): 209-220.
José de Paggi, S., 2001a . A new species of Lepadella (Rotifera: Monogononta: Colurellidae) from the Rio Pilcomayo National Park, Argentina. Hydrobiologia. 455: 223-228
José de Paggi, S., 2001b. Diversity of Rotifera (Monogononta) in wetlands of Río Pilcomayo National Park, Ramsar Site (Formosa, Argentina). Hydrobiologia 462: 25-34.
José de Paggi, S., 2002. New data on the distribution of Kellicottia bostoniensis (Rousselet, 1908) (Rotifera: Monogononta: Brachionidae): its presence in Argentina. Zool. anz. 241 (4): 363-368.
Kalff, J., 2002. Limnology . Prentice Hall, 592 p.
Koste, W. & Shiel, R., 1989. Classical taxonomy and modern methodology. Hydrobiologia 186/187: 279-284.
Marinone, M.C., 1994. A new and phylogenetically suggestive morphotype of Keratella lenz i (Rotifer, Monogononta), from Argentina. Hydrobiologia 299: 249-257.
Menge, B. & Sutherland, J.P., 1976. Species diversity gradients: synthesis of the roles of predation, competition, and temporal heterogeneity. Am. Nat. 110: 351-369.
Modenutti, B. & Claps, M.C., 1988. Monogononta Rotifers from plankton and periphyton of Pampasic lotic environments. Limnologica 19: 167-175.
Neiff, J.J., 1990. Ideas para la interpretación ecológica del Paraná. Interciencia 15: 441.
Nielsen, D.L., Shiel, R.J. & Smith,F.J.,1998. Ecology versus taxonomy: is there a middle ground? Hydrobiologia 387/388: 451-457.
Nogrady, T., Wallace, R. & Snell, T., 1993. Rotifera. Biology, Ecology and Systematics. 142 p. En Dumont, H. (Ed) Guides to the Identification of the Microinvertebrates of the Continental Waters of the World. SPB. Academic Publishing bv.
Prance, G.T., 1995. Systematics, conservation and sustainable development. Biodiversity and. Conservation 4: 490-500.
Quicke, D.L.J., 1993. Principles and tecniques of contemporary taxonomy. Blackie Academic & Professional. 311 p.
Ricklefs, R.E., 1998. Invitación a la Ecología. La economía de la naturaleza . Panamericana. 692 p.
Ruttner- Kolisko, A., 1989. Problems in taxonomy of rotifers, exemplified by the Filinia longiseta-terminalis complex. Hydrobiologia 186/187: 291-298.
Segers, H., 2002. The nomenclature of the Rotifera: annotated checklist of valid family and genus group names. J. Nat. Hist. 36: 631-640.
Segers, H., Nwadiaro, C.S. & Dumont, H., 1993. Rotifera of some lakes in the floodplain of the River Niger(Imo State Nigeria). II Faunal composition and diversity. Hydrobiologia 250: 63-71.
Welch, J.L.M. & Meselson, M., 1998. Karyotypes of bdelloid rotifers from three families. Hydrobiologia 387/388: 403-407.
Wheeler, Q.D. 1995. Systematics, the scientific basis for inventories of biodiversity. Biodiversity and Conservation 4: 476-489.
Fig. 1. Porcentaje de participación de algunos géneros en la composición de rotíferos Monogononta del Río Paraná
Subclase Monogononta Plate, 1889
Orden Ploima Hudson and Gosse, 1886
- Asplanchnidae Eckstein, 1883
Asplanchna Gosse, 1850
Asplanchnopus Guerne, 1888
- Brachionidae Ehrenberg, 1838
Anuraeopsis Lauterborn, 1900
Brachionus Pallas, 1766
Kellicottia Ahlstrom, 1938
Keratella Bory de St. Vincent, 1822
Notholca Gosse, 1886
Plationus Segers, Murugan and Dumont, 1993
Platyias Harring, 1913
- Dicranophoridae Harring, 1913
Dicranophoroides De Smet, 1997
Dicranophorus Nitzsch, 1827
- Epiphanidae Harring, 1913
Epiphanes Ehrenberg, 1832
- Euchlanidae Ehrenberg, 1838
Beauchampiella Remane, 1929
Dipleuchlanis De Beauchamp, 1910
Euchlanis Ehrenberg, 1832 (14 spp.)
- Gastropodidae Harring, 1913
Ascomorpha Perty, 1850
Gastropus Imhof, 1888
- Lecanidae Remane, 1933
Lecane Nitzsch, 1827
- Lepadellidae Harring, 1913
Colurella Bory de St. Vincent, 1824
Lepadella Bory de St. Vincent, 1826
Squatinella Bory de St. Vincent, 1826
- Mytilinidae Harring, 1913
Lophocharis Ehrenberg, 1838
Mytilina Bory de St. Vincent, 1826
- Notommatidae Hudson and Gosse, 1886
Cephalodella Bory de St. Vincent, 1826
Monommata Bartsch, 1870
Notommata Ehrenberg, 1830
- Scaridiidae Manfredi, 1927
Scaridium Ehrenberg, 1830
- Synchaetidae Hudson and Gosse, 1886
Ploesoma Herrick, 1885
Polyarthra Ehrenberg, 1834
Synchaeta Ehrenberg, 1832
- Trichocercidae Harring, 1913
Trichocerca Lamarck, 1801
- Trichotriidae Harring, 1913
Macrochaetus Perty, 1850
Trichotria Bory de St. Vincent, 1827
Wolga Skorikov, 1903
Orden Flosculariaceae Harring, 1913
- Conochilidae Harring, 1913
Conochilus Ehrenberg, 1834
- Filiniidae Harring and Myers, 1926
Filinia Bory de St. Vincent, 1824
- Flosculariidae Ehrenberg, 1838
Ptygura Ehrenberg, 1832
Sinantherina Bory de St. Vincent, 1826
- Hexarthridae Bartos, 1959
Hexarthra Schmarda, 1854
- Testudinellidae Harring, 1913
Pompholyx Gosse, 1851
Testudinella Bory de St. Vincent, 1826
- Trochosphaeridae Harring, 1913
Horaella Donner, 1949
Trochosphaera Semper, 1872
Order Collothecaceae Harring, 1913 (37)
- Collothecidae Harring, 1913
Collotheca Harring, 1913
Tabla 1. Familias y géneros de Rotíferos registrados en el río Paraná (ambientes lóticos y lénticos) desde su confluencia con el río Paraguay a la desembocadura en el río de la Plata.

Fig.2. Total de especies descriptas para distintos géneros de Rotíferos Monogononta y total de especies registradas para el río Paraná (en negro).
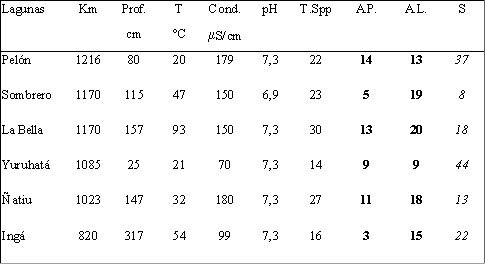
Tabla 2. Riqueza total de rotíferos (T.Spp), riqueza en el área pelágica (A.P.) y litoral (A.L.) en lagunas de la llanura de inundación del Paraná, ubicadas por kilómetros desde el Pto. de Buenos Aires. Se señala la afinidad faunística ( S) entre las áreas pelágicas y litoral de cada laguna