Las Ingresiones marinas del Neógeno en el sur de Entre
Ríos (Argentina) y Litoral Oeste de Uruguay y su
contenido malacológico.
Sergio MARTINEZ1 y Claudia DEL RÍO2
1Facultad de Ciencias- Departamento de Evolución de Cuencas - Iguá 4225. (11400) Montevideo - Uruguay.
E-mail: smart@fcien.edu.uy
2Museo Argentino de Ciencias Naturales Bernardino Rivadavia - División Paleoinvertebrados. Angel Gallardo
470 (C1405DJR) Buenos Aires-Argentina. E-mail: cdelrio@macn.gov.ar
Abstract: MOLLUSC FAUNAS OF THE NEOGENE MARINE TRANSGRESSIONS IN SOUTHERN ENTRE RIOS AND WESTERN LITTORAL OF URUGUAY.- Positive oscilations of the sea level during the Late Miocene, Late Pleistocene, and Holocene deposited fossiliferous sequences exposed along Argentinian and Uruguayan littorals. The study of the molluscan assemblages allowed to make inferences about some environmental factors, mainly salinity and temperature. According to faunistic evidence, the relatively high values of species richness, salinity and temperature recorded at the Late Miocene (corresponding to warm and normal salinity conditions), were progressively decreasing during subsequent transgressions. However, even in the youngest deposits (Holocene), the taxonomic composition indicates higher salinity and temperature values than at present. Since around 3000 years AP, there are no records of estuarine or marine molluscs in the studied area.
Key words: Molluscs - Neógeno transgretion - Argentina - Uruguay.
Palabras clave: Moluscos - Transgresión Neógeno - Argentina - Uruguay.
Introducción
Los diversos procesos geológicos que afectaron la región mesopotámica argentina han modelado su geomorfología e influido en forma importante en la composición y distribución de la biota que la caracteriza, siendo las transgresiones marinas de los últimos millones de años las que tuvieron una de las mayores influencias en este aspecto. La presente contribución es una síntesis de los datos que se poseen sobre las faunas de moluscos de las tres invasiones marinas del Neógeno (en su sentido más actual, ver International Commission on Stratigraphy, 2004) registradas en el litoral oeste de Uruguay y en el sur de Entre Ríos (Argentina). El contenido malacológico registrado en los depósitos dejados por el mar ha permitido en todos los casos inferir la salinidad y temperaturas relativas de sus aguas, y en el caso de los registros más modernos, interpretar sus edades radiocarbónicas.
Los datos de base proceden de trabajos previos, los que serán citados oportunamente, con la excepción de los referidos a los del Holoceno de Uruguay, que son en su mayoría novedosos (Martínez et al., inédito). Toda esta información se enmarca en un contexto detallado de gradientes de salinidad y diversidad taxonómica regional mediante la comparación de las asociaciones neógenas uruguayo-mesopotámicas con las existentes en los ambientes actuales del Río de la Plata y su frente marítimo, confirmándose una vez la utilidad de los moluscos como herramientas paleoecológicas.
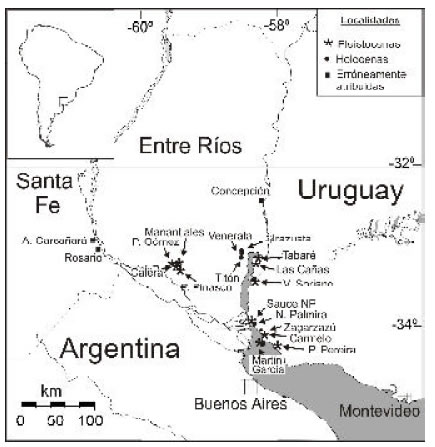
Las localidades del Pleistoceno y Holoceno mencionadas en el texto se indican en la figura 1, y las miocenas se encuentran en detalle en Del Río y Martínez (1988a).
Fig 1. Ubicación de las localidades cuaternarias mencionadas en el texto.
La transgresión del Mioceno Tardío
Un punto de inflexión en el modelo paleocirculatorio del Atlántico Sur lo constituyó la apertura del pasaje de Drake y el subsecuente desarrollo de la Corriente Circumpolar Antártica (Shackleton y Kennet 1975a, b, Kennet 1977, 1980, Scasso y Castro, 1999, Scasso et al., 2000, Barker y Thomas, 2004). La influencia de su derivada Corriente de Malvinas se hizo sentir sobre las faunas de moluscos recién a partir de finales del Mioceno (Martínez y Del Río, 2002 a y b), momento durante el cual una gran extensión del territorio de Argentina y Uruguay fueron ocupados por el «mar entrerriense o paranense». Los depósitos dejados por esta transgresión marina hace ca. 10 MA (Zinsmeister et al., 1981; Scasso et al. 2001) corresponden a las formaciones Camacho (Uruguay), y Paraná y Puerto Madryn (Argentina), portadores de una típica asociación de moluscos estudiados detalladamente en sus aspectos sistemáticos por Del Río (1991, 1992, 1994), Martínez (1994), Del Río y Martínez (1998 a, b), Martínez et al. (1998).
Martínez y Del Río (2002 a) propusieron para la región tratada la existencia durante el Mioceno Tardío de la Bioprovincia Paraniana (Fig. 2) la que abarcaba las asociaciones de moluscos del sudoeste de Entre Ríos y del sur de Uruguay (ubicadas dentro de las formaciones Paraná y Camacho respectivamente). Como fue discutido en detalle por Martínez y Del Río (opp. cit.), la misma limitaba por el sur con la Provincia Valdesiana y siempre dentro del espectro climático subtropical (paratropical sensu Petuch 1988), las aguas eran más cálidas que en esta última y que las actuales en las mismas latitudes, caracterizándose por la presencia de elementos eutropicales (Strombus y Terebra) y por un alto porcentaje de taxones paratropicales. La existencia de aguas cálidas respondió a los movimientos tectónicos que siguieron a la apertura del pasaje de Drake, los que seguramente desviaron hacia el este a la Corriente de Malvinas o proto Malvinas, por aquel entonces de influencia incipiente. Esta malacofauna refleja oscilaciones de la salinidad entre valores normales, indicados por la presencia de equinodermos y braquiópodos asociados, a polihalinos al menos, como los inferidos por el desarrollo de biostromas ostreros o a través de las asocaciones foraminiferalógicas (Del Río y Martínez, 1998 a, Aceñolaza, 2000, Marengo, 2000).
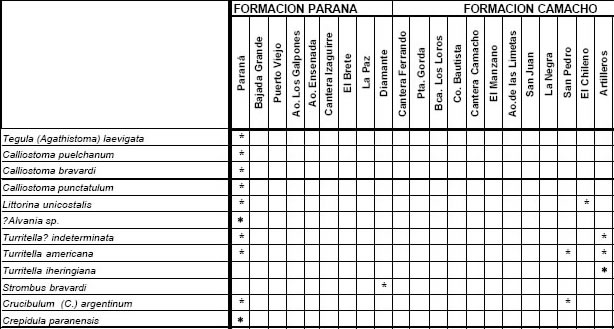
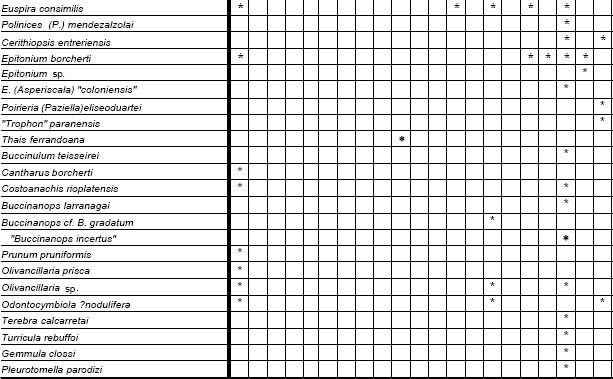
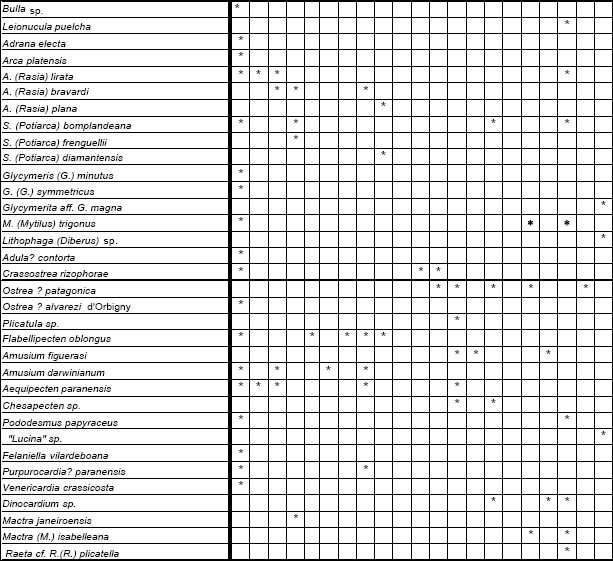
Las 86 especies recuperadas de esta unidad (Fig. 3) corresponden a 53 taxones procedentes del margen derecho del Río Paraná y 49 recuperados del sudoeste del Uruguay, localidades que, con excepción de Los Galpones , y de acuerdo con su composición taxonómica se reúnen claramente en un grupo occidental (Entre Ríos) y otro oriental (Uruguay) (Fig. 4), reflejando probablemente diferentes factores ambientales que dominaron ambas áreas geográficas.
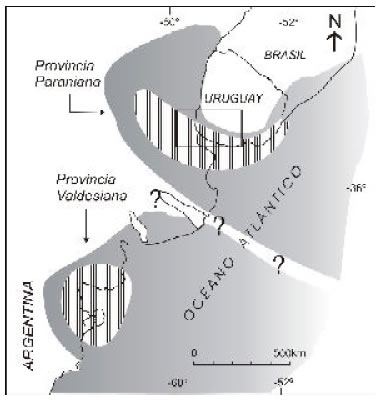
Fig 2. Las bioprovincias Valdesiana y Paraniana del Mioceno Tardío. El rectángulo señala el área objeto de este trabajo.
 |
 |
 |
 |
Fig 3. Especies de la Provincia Paraniana (por localidades), Mioceno.
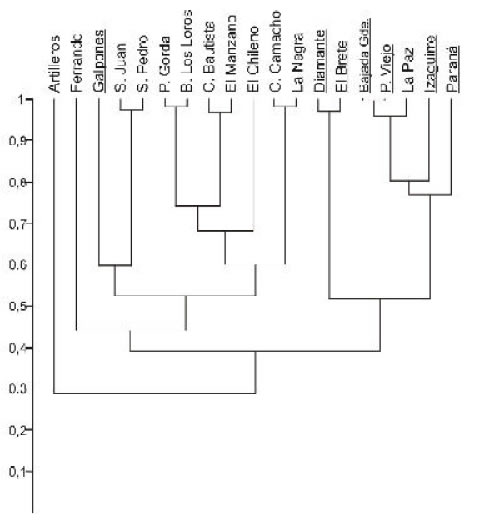
Fig 4. Agrupamiento (UPGMA, índice de Raup & Crick) de localidades de la Bioprovincia Paraniana. Se indican subrayadas las argentinas.
La trangresión del Pleistoceno Tardío
Por lo menos desde que Ameghino (1889) acuñó el término “Belgranenese”, se reconoce una transgresión pleistocena que cubrió parte de la provincia de Buenos Aires. Desde ese entonces, y habiéndose también atribuido a ella depósitos allende esta provincia, una larga sucesión de términos y atribuciones temporales se fue dando a lo largo de los años (ver síntesis en Aguirre y Whatley, 1995, Isla et al. 2000, Aguirre y Fucks, 2004, Martínez y Ubilla, 2004), discusión que excede los propósitos de este trabajo, por lo que nos limitaremos a hablar genéricamente de “depósitos pleistocenos”.
En el área que estamos tratando, estos depósitos se han encontrado en la costa uruguaya (Martínez et al. 2001, Rojas 2004), en la Isla Martín García (González y Ravizza, 1987) y en la región entrerriana situada al sur de Gualeguaychú (Guida y González, 1984).
Las edades 14C adjudicadas a todos estos depósitos en base a los moluscos se pueden consultar en la figura 5, donde se aprecia que prácticamente todas ellas se encuentran en las cercanías del límite de resolución del método, por lo que deben ser consideradas edades mínimas. En el caso de las localidades uruguayas, y como se explicitará más adelante, de acuerdo con la temperatura inferida, Martínez et al. (2001) propusieron que estos moluscos vivieron durante el piso isotópico 5e (último interglacial, hace alrededor de 120 000 años). En cambio, la pobreza taxonómica de las asociaciones entrerrianas y de Martín García no permite la obtención de información adecuada sobre la paleotemperaturas en esas localidades, pero dada la cercanía geográfica, es razonable suponer la misma edad que para las uruguayas. La misma edad ha sido atribuida también por Isla et al. (2000) a los depósitos “Belgraneneses” de la Provincia de Buenos Aires, basándose en inferencias a partir de dataciones uranio/torio.
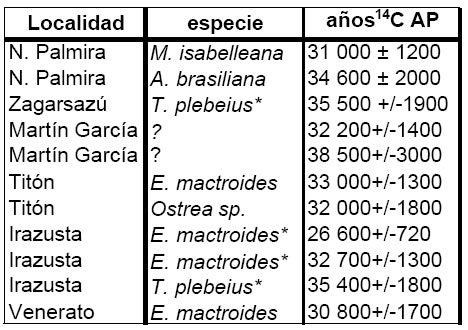
Fig 5. Edades 14C para las asociaciones pleistocenas. El asterisco indica ejemplares colectados en posición de vida. Datos de Guida y González (1984), González y Ravizza (1987), Martínez et al. (2001) y Rojas (2004).
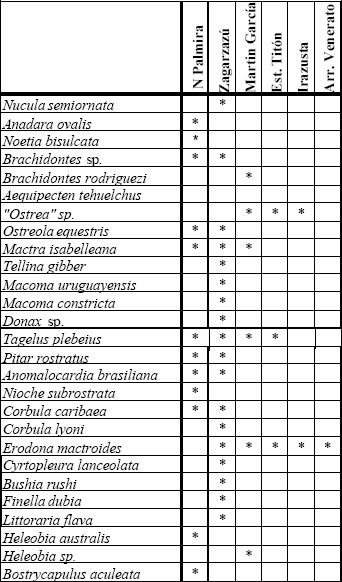
Los taxones considerados para las comparaciones de este trabajo (37 especies) pueden consultarse en la figura 6, confeccionada sobre la base de los datos proporcionados por Guida y González (1984), González y Ravizza (1987), Martínez et al. (2001) y Rojas (2004).
 |
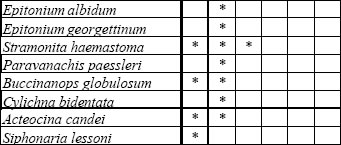 |
Fig 6. Especies pleistocenas, por localidades. Fuentes igual que en la figura 5.
La riqueza específica presenta un claro gradiente decreciente en sentido este-oeste (Fig. 7), vinculado con la disminución de la salinidad en esa dirección, tal como lo indican los porcentajes de especies con diferente tolerancia salina, y emblemáticamente la distribución de la típica especie estuarina (oligohalina) Erodona mactroides Daudin in Bosc, 1801. Mientras esta especie solo está presente en una localidad uruguaya, donde está asociada a otras de salinidad normal, se constituye en componente importante de las asociaciones entrerrianas y de Martín García, siendo inclusive la única especie presente en la localidad de Venerato (Entre Ríos). Un comentario aparte merece la asociación de Martín García, donde podría esperarse una mayor riqueza específica teniendo en cuenta su ubicación relativa en el referido gradiente salino; en este sentido, puede que dicho fenómeno responda a un muestreo menos intenso.
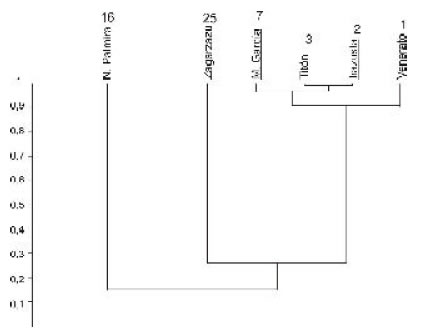
Fig 7. Agrupamiento (WPGMA, índice de Raup & Crick) de las localidades pleistocenas. Se indican subrayadas las argentinas. Los números junto al nombre de la localidad corresponden a la cantidad de especies.
Respecto a la temperatura de las aguas, y en base tanto al registro de especies con diferentes afinidades térmicas como a la presencia de algunos taxones que hoy están restringidos a latitudes más septentrionales que durante el Pleistoceno [Nioche subrostrata (Lamarck, 1818), Anomalocardia brasiliana (Gmelin, 1791), Macoma constricta (Bruguière, 1792) y Littoraria flava (King & Broderip, 1832)], Martínez et al. (2001) postularon para ese momento una temperatura mayor a la actual en el litoral uruguayo, que se correspondería con la alcanzada durante el transcurso del piso isotópico 5e, cuando se llegó a un registro térmico mayor al actual (Krantz et al., 1987, Zubakov y Borzenkova, 1990; Murray-Wallace y Belperio, 1991).
Debido a la muy baja riqueza taxonómica del litoral entrerriano y de Martín García, no es posible realizar para estas localidades cálculos fiables basándose en las preferencias térmicas de las especies. No se encuentran en los referidos depósitos especies “termofílicas” como las señaladas en el párrafo anterior para Uruguay, aunque por lo menos para el caso de Entre Ríos, la baja salinidad de las aguas habría impedido de todos modos su existencia.
La transgresión del Holoceno
Al igual que para el Pleistoceno, los registros de la transgresión Holocena se conocen desde larga data, aunque la diferenciación entre ambas estrictamente por su contenido fosilífero ha sido problemática. Las dataciones radiocarbónicas han permitido separar estos dos episodios, por lo que trataremos aquí únicamente aquellos depósitos holocenos fechados en forma “absoluta”, como los estudiados por Guida y González (1984), González y Gavizza (1987) y Martínez et al. (inédito). La lista de especies analizadas (24) puede consultarse en la figura 8, y las edades 14C y AP calculadas en la Fig. 9. Como se observa en dicha figura, existe un registro de ca. 10 000 años cal. AP en Sauce de Nueva Palmira (un fechado muy antiguo y extraño para el Atlántico Sudoccidental), y luego una serie de edades más o menos continua a partir de los ca. 6000-6500 años cal. AP y hasta alrededor de los 3 000 años cal. AP.
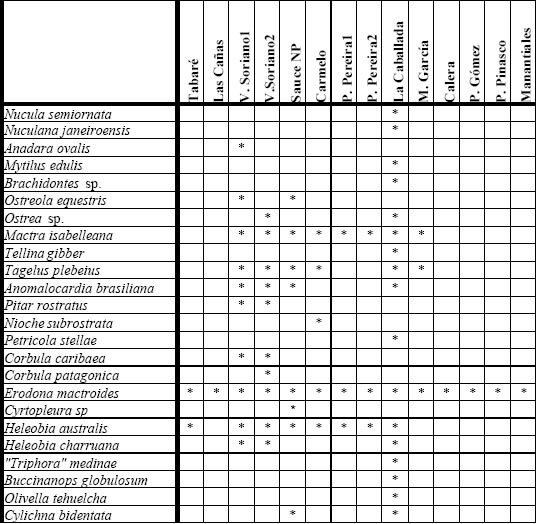
Fig 8. Especies del Holoceno, por localidades. Datos de Guida y González (1984), González y Gavizza (1987) y Martínez et al. (inédito).
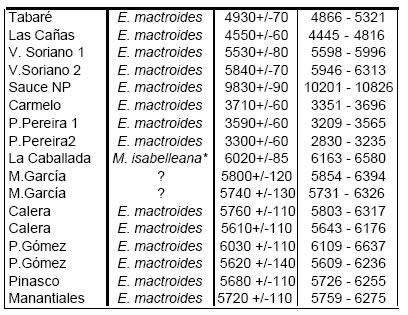
Fig 9. Edades 14C y años calendario antes del presente (1950 por convención) calculadas de acuerdo con Stuiver y Reimer (1993), para las asociaciones holocenas. El asterisco indica ejemplares colectados en posición de vida. Las edades calculadas de los moluscos uruguayos se indican según Martínez et al. (inédito), y las argentinas a partir de las edades 14C brindadas por Guida y González (1984) y González y Ravizza (1987).
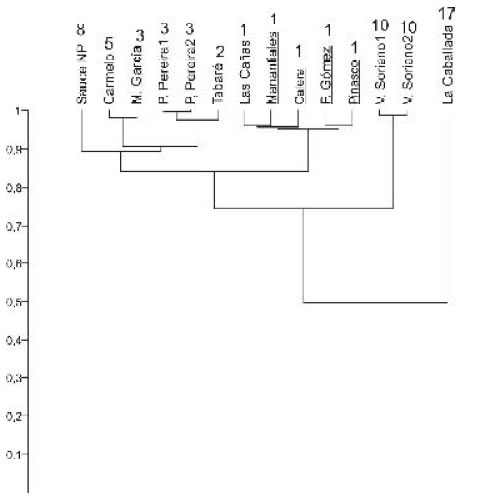
Fig 10. Agrupamiento (UPGMA, índice de Raup & Crick) de las localidades holocenas. Se indican subrayadas las argentinas. Los números junto al nombre de la localidad corresponden a la cantidad de especies.
Como ya fue indicado por Guida y González (1984), en las localidades entrerrianas los registros se limitan a la presencia de Erodona mactroides indicando claramente la existencia de aguas salobres. En la costa uruguaya, esta especie también se encuentra presente, pero ya formando parte de asociaciones más diversas (en especial en La Caballada) y con especies que requieren una mayor salinidad, indicando un aumento en este parámetro hacia el este.
Ateniéndonos a la presencia en el Holoceno de la referida Erodona mactroides en áreas donde estuvo ausente durante el Pleistoceno, y también a la mayor riqueza específica existente en las asociaciones de este último lapso, la salinidad debe de haber sido menor en la región considerada durante la transgresión holocena.
Las asociaciones entrerrianas, cuando comparadas con las uruguayas (figura 10), se aglomeran en primera instancia con Las Cañas, donde también E. mactroides es la única especie, y luego con las más pobres taxonómicamente. Martín García, de acuerdo con su posición geográfica, se vincula claramente con asociaciones de la costa oriental. La Caballada, que permanece aparte del conjunto, representa el depósito más diverso en cuanto a número de especies. Otro hecho a destacar es que Sauce de Nueva Palmira no se separa taxonómicamente del resto de las asociaciones, a pesar de su edad claramente más antigua (ver más arriba).
La composición de las asociaciones uruguayas permite inferir las temperaturas de las aguas holocenas. La evidencia de marcas térmicas mayores a las actuales es constante a lo largo de todos los afloramientos, y queda demostrada por la presencia de indicadores de aguas cálidas como los son Anomalocardia brasiliana y Nioche subrostrata, especies que hoy están ausentes en estas mismas latitudes, y por la proporción de especies que llegan a la región por influencia de la corriente cálida de Brasil. De todos modos, no se habrían alcanzado los registros térmicos del Pleistoceno.
Al igual que en el caso de los depósitos pleistocenos, Martín García y las localidades entrerrianas cuentan con muy pocas especies (una sola en el segundo caso), lo que hace imposible una estimación de la paleotemperatura por afinidades térmicas de las mismas.
En síntesis
El litoral argentino y su inmediato uruguayo fueron afectados por tres eventos de aumento del nivel del mar que dejaron testigos malacológicos. Estos corresponden al Mioceno Tardío, Pleistoceno Tardío, y Holoceno. La riqueza de especies fue disminuyendo progresivamente, en consonancia con las propias evidencias faunísiticas de una menor temperatura y salinidad en cada oportunidad. Empero, aún en el caso más cercano (Holoceno), la composición taxonómica indica una temperatura (y obviamente, salinidad) mayor a la presente. A partir de alrededor de los 3000 años AP antes del presente, no se han registrado moluscos marinos o estuarinos en el área considerada.
Apéndice. Referencias dudosas a moluscos marinos cuaternarios
1) Ihering (1907:428) mencionó que le fue entregada una pequeña colección de moluscos cuaternarios procedente del Río Uruguay, cerca de Concepción del Uruguay, indicando la presencia de (nomenclatura y autores textual): Bulla striata Brug., Siphonaria Lessoni Blv., Acmaea subrugosa Orb., Cerithium atratum Born., Urosalpix Rushi Pils., Pododesmus rudis Brod., Phacoides pectinatus Gm., Chione cancellata L., Chione portesiana Orb., Anomalocardia brasiliana Gm.. Resulta muy extraño que estas especies llegaran tan al norte durante cualquiera de las ingresiones cuaternarias, teniendo en cuenta lo expresado más arriba respecto a las salinidades, tanto en Uruguay como en Argentina. Doello Jurado (1940) y Parodiz (1962) consideraron que B. striata, C. atratum, P. rudis, P. pectinatus, C. cancellata, y C. portesiana, eran parte del balastro descargado por barcos brasileños y que S. lessoni, A. subrugosa, U. rushii y A. brasiliana provendrían de “depósitos pleistocénicos” de Gualeguaychú. De todos modos, sigue siendo muy extraña la supuesta presencia de estas especies en Gualeguaychú, a la vista de las investigaciones posteriores.
2) Castellanos (1943), mencionó afloramientos con moldes de Corbula mactroides, Mactra isabelleana, Anomalocardia brasiliana, Tagelus gibbus y Panopaea sp. (nomenclatura textual) en los alrededores de Rosario, correspondientes según este autor al “Interensenadense” y al “Belgranense”. El escaso material conservado en el Instituto de Fisiografía y Geología de Rosario no permite realizar con seguridad asignaciones específicas, pudiéndose tratar inclusive de especies dulciacuícolas. Algunas conchillas mal conservadas, con procedencia en el margen izquierda del Arroyo Carcañará a unos 3 km de su desembocadura, sí se asemejan mucho a Erodona mactroides, pero serán necesarias nuevas colectas para corroborarlo y para precisar su situación estratigráfica (“Belgranense” según Castellanos, op.cit.).
3) Prost (1982, basada en Ecochard (1970, inédito) mencionó la presencia de moluscos marinos o de agua salobre para la localidad de Itapebí (aproximadamente 20 km al norte de la ciudad de Salto, Uruguay), sugiriendo que las sedimentitas portadoras correspondían a la transgresión Flandriana/ Dunkerkiana? (Formación Villa Soriano en Uruguay, según la autora). Representantes de los géneros mencionados en ese trabajo (sic: Neocorbicula, Diplodon, Potamolithus, Littoridina), con la excepción de Littoridina (Heleobia Stimpson, 1865, de acuerdo con la reubicación de las especies de la región), fueron hallados efectivamente allí en otra oportunidad (ver Martínez, 1988), pero se trata de organismos de agua dulce.
Agradecimientos
Horacio Parent facilitó amablemente el acceso a la colección A. Castellanos alojada en el Instituto de Fisiografía y Geología de la Universidad Nacional de Rosario.
Bibliografía
Aceñolaza F. G. 2000. La Formación Paraná (Mioceno medio): estratigrafía, distribución regional y unidades equivalentes. En, F. G. Aceñolaza y R. Herbst (eds.), El Neógeno de Argentina. Serie Correlación Geológica, 14: 9- 27
Aguirre, M.L. y Fucks, E. 2004. Moluscos y paleoambientes del Cuaternario marino en el sur de Entre Ríos y litoral bonaerense. En, F. G. Aceñolaza (ed.), Temas de la Biodiversdiad del litoral fluvial argentino, INSUGEO, Miscelánea 12: 55-70.
Aguirre, M. L., and Whatley, R. C. (1995). Late Quaternary marginal marine deposits and palaeonvironments from northeastern Buenos Aires Province, Argentina: a review. Quaternary Science Reviews 14: 223-254.
Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias de Córdoba, 6: 1-1027
Barker, P.F. y , E. Thomas. 2004. Origin, signature and palaeoclimatic influence of the Antarctic Circumpolar Current. Earth-Science Reviews 66 : 143–162.
Castellanos, A. 1943. El Preensenadense es un horizonte geológico o una facies?. Publicaciones del Instituto de Fisiografía y Geología, 18: 1-84. Rosario
del Río, C. J. 1991. Revisión sistemática de los Bivalvos de la Formación Paraná (Mioceno Medio), Provincia de Entre Ríos - Argentina. Academia Nacional de Ciencias Exactas, Físicas y Naturales, Monografías 7: 11-93. Buenos Aires.
del Río, C. J. 1992. Middle Miocene Bivalves of the Puerto Madryn Formation, Valdés Peninsule, Chubut Province, Argentina. (Nuculidae-Pectinidae), Part I: Palaeontographica (A) 225, 1-57. Sttutgart.
del Río, C. J. 1994. Middle Miocene Bivalves of the Puerto Madryn Formation, Valdés Peninsule, Chubut Province, Argentina. (Lucinidae - Pholadidae), Part II: Palaeontographica (A) 231, 93-132. Stuttgart.
del Río, C. J. y Martínez, S., 1998a, El Mioceno marino en la Argentina y en el Uruguay: En, C. J. del Río (ed.), Moluscos marinos Miocenos de la Argentina y del Uruguay. Academia Nacional de Ciencias Exactas, Físicas y Naturales, Monografías 15: 6-25. Buenos Aires.
del Río, C. J. y Martínez, S., 1998b. Clase Bivalvia: En, C. J. del Río (ed.), Moluscos marinos Miocenos de la Argentina y del Uruguay. Academia Nacional de Ciencias Exactas, Físicas y Naturales, Monografías 15: 48-83. Buenos Aires.
Martínez, S., Scasso, R. A., 2001. Nature and origin of spectacular marine Miocene shell beds of northeastern Patagonia (Argentina): Paleoecological and bathymetric significance. Palaios 16, 3-25.
Doello Jurado, M. 1940. Presencia de Moluscos Marinos en los Yacimientos Arqueológicos de Santiago del Estero. Síntesis Malacológica, 2: 123-144.
González, M.A. y Ravizza, G. 1987. Sedimentos estuáricos del Pleistoceno Tardío y Holoceno en la isla Martín García, Río de la Plata. Revista de la Asociación Geológica Argentina, 42 (3-4): 231-243.
Guida, N.G. y González, M.A. 1984. Evidencias paleoestuáricas en el sudeste de Entre Ríos, su evolución con niveles marinos relativamente elevados del Pleistoceno Superior y Holoceno. 9º Congreso Geológico Argentino (S.C. de Bariloche, Río Negro), Actas 3: 577-594. Buenos Aires.
Ihering, H. von., 1907. Les Mollusques fossiles du Tertiaire et du Cretace superieur de l’ Argentine. Anales del Museo Nacional (3) 7, 1-611. Buenos Aires.
Isla, F.I., Rutter, N.W., Schnack, E.J. y Zárate, M.A. 2000. La transgresión Belgranense en Buenos Aires. Una revisión a cien años de su definición. Revista Cuaternario y Ciencias Ambientales, Publicación Especial, 4: 3-14.
Kennet, J. P., 1977. Cenozoic evolution and Antarctic glaciation, the Circum-Antarctic Ocean an their impact on global Paleoceanography. Journal of Geophysical Research, 82, 3843-3860.
Kennet, J. P., 1980. Paleoceanographic and biogeographic evolution of the Southern ocean during the Cenozoic and Cenozoic microfossils datum. Palaeogeography, Palaeoclimatology, Palaeoecology, 31, 123-152. Amsterdam.
Krantz, D. E., Williams, D. F., y Jones, D. S. 1987. Ecological and paleoenvironmental information using stable isotope profiles from living and fossil molluscs. Palaeogeography, Palaeoclimatology, Palaeoecology, 58: 249-266. Amsterdam.
Marengo H. G. 2000. Rasgos micropaleontológicos de los depósitos de la transgresión Entrerriense-Paranense en la cuenca Chaco-Paranense y Noroeste Argentino. . En, F. G. Aceñolaza y R. Herbst (eds.), El Neógeno de Argentina. Serie Correlación Geológica, 14: 29-45
Martínez, S., 1988. Adiciones a la malacofauna de la Fm. Sopas (Pleistoceno Superior, Uruguay), con algunas consideraciones paleoecológicas. Paulacoutiana 3: 67-75.
1994. Bioestratigrafía (invertebrados) de la Formación Camacho (Mioceno, Uruguay). Tesis de Doctorado. Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Argentina.
Martínez, S., y del Río, C. J. . 2002. Late Miocene Molluscs From the Southwestern Atlantic Ocean (Argentina and Uruguay): A Palaeobiogeographic Analysis. Palaeogeography, Palaeoclimatology, Palaeoecology, 188 (3-4): 167-187. Amsterdam.
Martínez, S., y del Río, C. J. 2002. Las Provincias Malacológicas Miocenas y Recientes del Atlántico Sudoccidental. Anales de Biología, 24: 121-130 Murcia.
Martínez, S., y Ubilla, M. 2004. El Cuaternario de Uruguay . En, G.Veroslavsky, M. Ubilla y S. Martínez, Cuencas sedimentarias de Uruguay. Cenozoico: 195-228. EUDECI. Montevideo.
Martínez, S., del Río, C. J., Reichler, V. 1998. Clase Gastropoda: En, C. J. del Río (ed.), Moluscos marinos Miocenos de la Argentina y del Uruguay. Academia Nacional de Ciencias Exactas, Físicas y Naturales, Monografías 15, pp. 26- 47. Buenos Aires.
Martínez, S., Ubilla, M., Verde, M., Perea, D., Rojas, A., Guérèquiz, R. and Piñeiro, G. 2001. Paleoecology and Geochronology of Uruguayan Coastal Marine Pleistocene Deposits. Quaternary Research 55: 246-254.
Martínez, S., Rojas A, Ubilla M, Verde M, Perea D, Guérequiz R. y Piñeiro G (inédito): Shell-beds from the marine Holocene of Uruguay: Geochronology and Paleoecology. enviado para su publicación a Ameghiniana.
Murray-Wallace, C. V. y Belperio, A. P. 1991. The last Interglacial Shoreline in Australia – A Review. Quaternary Science Reviews 10: 441-461.
Parodiz, J.J. 1962. Los moluscos marinos del Pleistoceno rioplantense. Comunicaciones de la Sociedad Malacológica del Uruguay, 1(2): 29-46.
Prost, M.T., 1982. Heritages Quaternaires et évolution géomorphologique des bords du Rio de la Plata en Uruguay. Tesis de Doctorado Université de Paris VII. 597 pp.
Rojas, A. 2004. La asociación de moluscos fósiles de Zagarzazú, Pleistoceno Tardío, Uruguay. 4º Congreso Uruguayo de Geología (Montevideo), Actas (CdRom), Contribución 26, 3pp. Uruguay.
Scasso, R.A. y Castro, L.N. 1999. Cenozoic phosphatic deposits in North Patagonia, Argentina: Phosphogenesis, sequence-stratigraphy and paleoceanography. Journal of South American Earth Sciences, 12: 471-487.
Castro, L. N. y Tofalo, O. R., 2000. Phosphogenesis, sequence-stratigraphy and paleoceanography in Gaiman Formation phosphates, Argentina. 31st International Geological Congress (Río de Janeiro). Abstract 2344 (Cd Rom).
McArthur J. M., del Río, C. J., Martínez, S. y Thirlwall M.F., 2001. 87Sr/86Sr Late Miocene age of fossil molluscs in the ”Entrerriense” of the Valdés Península (Chubut, Argentina). Journal of South American Earth Sciences, 14: 319-327.
Shackleton, N. y Kennet, J. P. 1975a. Paleotemperatures history of the Cenozoic and the initiation of Antarctic glaciation: oxygen and carbon isotopic analysis in DSDP 277, 279 and 281. Initial Reports of the Deep Sea Drilling Project, 29: 743.
Shackleton, N. y Kennet, J. P. 1975b. Late Cenozoic oxygen and carbon isotopic changes. DSDP Site 284: Implication for glacial history of northern Hemisphere and Antarctica. Initial Reports of the Deep Sea Drilling Project, 29: 801.
Stuiver, M. y Reimer, P. J. 1993. Extended 14C database and revised CALIB radiocarbon calibration program. Radiocarbon, 35: 215-230.
Zinsmeister, W. J., Marshall, L.G., Drake, R.E. y Curtis, G.H. 1981. First Radioisotope (Potassium-Argon) Age of marine Neogene Rionegro beds in northeastern Patagonia, Argentina. Science, 212 (4493): 440.
Zubakov, V., y Borzenkova, I. (1990). Global palaeoclimate of the Late Cenozoic. Elsevier, New York. 456 pp.