Estudio palinoestratigráfico del Oligoceno tardío-Mioceno en secciones de la costa patagónica y plataforma continental argentina
Viviana BARREDA 1 y Susana PALAMARCZUK 2
Abstract.- PALINOSTRATIGRAPHICAL STUDY OF THE LATE OLIGOCENE-MIOCENE IN SECTIONS OF PATAGONIAN SHORE AND CONTINENTAL PLATFORM OF ARGENTINA. The purpose of this study is to provide a preliminary palynostratigraphic correlation scheme based on integrated distributions of both, continental and marine palynomorphs from surface and subsurface sections (Upper Oligocene through Early-Middle Miocene) located along the Atlantic Patagonic Coast and in the Argentine Continental Shelf between latitudes 40º and 53º S. The age assignment is supported by concurrent information from known local and regional stratigraphic ranges of the species and by the presence of other diagnostic fossil groups. Four informal angiosperm pollen and dinocyst associations are proposed: M-M/R(Late Oligocene), C-T/L, G/C (Early Miocene) and T-B/H (late Early - Middle Miocene). The Lower association is characterized by the presence of Mutisiapollis viteauensis (Barreda) Barreda, Margocolporites tenuireticulatus Barreda and ?Reticulatosphaera actinocoronata (Benedek) Bujak and Matsuoka, and was recognized in Mazarredo and Punta Nava sections, southern San Jorge Gulf area, and in Aries x-1 well, Austral Basin, interval 530-350m. In Ra x-1 well, Colorado Basin, below depth 1790m (Elvira Formation), the association was not clearly recognized due to the scarcity of dinocysts, although the interval might represent the proposed M-M/R association.
At level 1930m (the lowermost fertile sample) the presence of Tuberculodinium vancampoae (Rossignol) Wall indicates an age not older than Upper Oligocene. The C-T/L association , characterized by Cyperaceaepollis neogenicus Krutzsch, Tricolpites trioblatus Mildenhall and Pocknall, and Lingulodinium hemicystum Mc Minn, was identified in Ra x-1 well, interval 1790-1650m (top of Elvira Formation and basal Barranca Final Formation); Chenque Formation, lower Chenque section, San Jorge Gulf Basin; top of Punta Nava and El Faro sections in the southern San Jorge Gulf area, in the lower part of Monte Entrada and Monte Observación sections, Austral Basin, and in Aries x-1 well, interval 350-260m, Austral Basin. The G/C association, characterized by Glencopollis ornatus Pocknall and Mildenhall, and Cannosphaeropsis utinensis Wetzel, was recognized in Ra x-1 well between 1630 and 1610m, the uppermost level herein considered. Chenque Formation (upper Chenque section), San Jorge Gulf Basin; upper part of Monte Entrada and Monte Observacion sections, Austral Basin, Santa Cruz Province, are also included in this association. The T-B/H association, defined by the presence of Tubulifloridites antipodica Cookson, Baumannipollis chubutensis Barreda, and acmé of Hystrychosphaeropsis obscura Habib, was recognized in Antena and Cañadón Baumann sections, Chenque Formation, Chubut Province. This study indicates that integrated palynological data are of considerable promise for further, more detailed biostratigraphic correlation.
Key words: Palynostratigraphy. Correlation. Late Oligocene-Miocene. Patagonia. Continental Shelf. Argentina.
Palabras clave: Palinoestratigrafía. Correlación. Oligoceno Tardío. Mioceno. Patagonia. Plataforma continental. Argentina.
Introducción
En este trabajo se presenta una síntesis de las investigaciones palinológicas realizadas por lasautoras en distintas secciones en el entorno del Mioceno a lo largo de la costa atlántica y de la plataforma continental argentina. La región que abarca el estudio comprende un conjunto de afloramientos y secciones de pozo cuya interpretación, especialmente con respecto a la edad de las transgresiones marinas involucradas, ha sido controvertida. El concepto original de "formación patagónica" utilizado para caracterizar estos depósitos en los de D`Orbigny fue seguido por autores posteriores, atribuyéndole a la misma asociaciones fósiles provenientes de diversas localidades y edades que variaron entre el Eoceno y el Mioceno, o aún Plioceno (Hatcher, 1897; Ortmann, 1900; Ihering, 1902; Wichmann, 1918; Windhausen, 1924; Frenguelli, 1929; Feruglio, 1949). Estudios subsiguientes perfeccionaron este esquema original mediante
el análisis de perfiles de detalle, definición de unidades formacionales, aporte de nuevos conocimientos paleontológicos y dataciones radimétricas (Camacho y Fernández, 1956; Camacho, 1974; Becker, 1964; Becker y Bertels, 1980; Bertels, 1970, 1975, 1978; Malumián, 1970, 1972; Malumián y Náñez, 1988, 1991, 1998; Náñez, 1990; Cione y Expósito, 1978; Parma, 1985, 1989; Bellosi, 1990, 1995; Legarreta y Uliana, 1994; Feagle, et al., 1995, entre otros). La palinología es una disciplina que comenzó a ser usada algo más modernamente para caracterizar estos depósitos en distintas localidades, sumando un nuevo aporte para las interpretaciones estratigráficas locales (Gamerro y Archangelsky, 1981; Quattrocchio et al., 1986; Guerstein, 1987, 1990a y b; Guerstein y Quattrocchio, 1988; Guerstein y Guler, 2000; Archangelsky, 1996).
Los estudios palinológicos consisten en el análisis de microfósiles de pared orgánica de origen continental y marino. Dentro del primer grupo se consideran las algas de agua dulce, simples o coloniales, restos de hongos (esporas, cuerpos fructíferos e hifas), esporas de briófitas y pteridófitas y granos de polen de gimnospermas y de angiospermas. Forman parte del espectro orgánico continental, además de los restos figurados que se mencionaron, fragmentos microscópicos de vegetales, principalmente leños y cutículas. El segundo grupo, palinomorfos de origen marino, consiste fundamentalmente en quistes de dinoflagelados además de algas simples o coloniales, acritarcos y capas internas de foraminíferos.
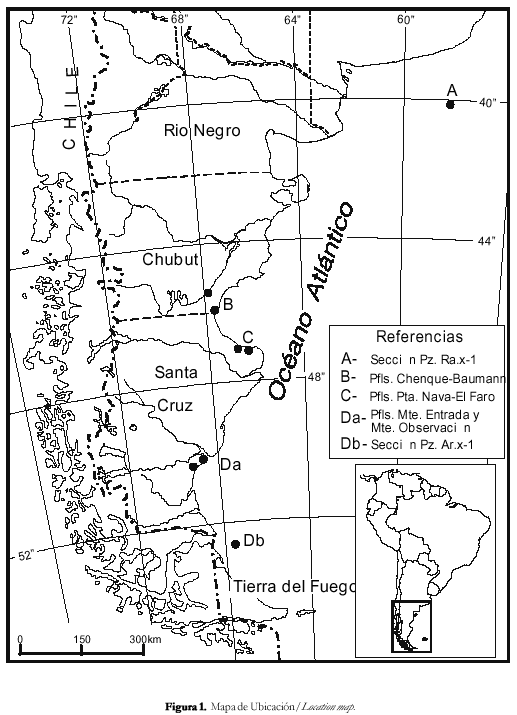
En este trabajo se intenta una correlación regional para un área latitudinalmente extendida, en base al análisis integrado de palinomorfos de origen continental y marino comparando secciones ubicadas entre los 40º y 53º S. Las localidades consideradas son: A- Cuenca del Colorado (pozo Ra x-1, profundidad 2030 a 1610mbbp, abarcando parte de la Fm. Elvira y la base de la Fm. Barranca Final, sólo paleomicroplancton), B- Cuenca del Golfo San Jorge, Fm. Chenque (perfiles Chenque, Viteau, Punta Delgada, Playa Las Cuevas, Antena y Baumann), C- Área sur del Golfo San Jorge (perfiles Mazarredo, Punta Nava y El Faro), D- Cuenca Austral, a- Formación Monte León (perfiles Monte Observación y Monte Entrada), b- Pozo Aries x-1 (profundidad 640- 260mbbp) (figura 1).
En cada una de las localidades estudiadas se procedió al estudio conjunto de los grupos considerados lográndose resultados de carácter cronoestratigráfico, y, en parte, paleoclimático y paleobiogeográfico. El análisis se llevó a cabo separadamente para cada uno de los grupos, aunque los mismos se recuperaron de manera conjunta a partir del tratamiento estándar para palinología. Este abordaje es metodológicamente adecuado debido a las diferencias resultantes del distinto origen y requerimientos ecológicos de cada uno.
En esta síntesis se presenta un esquema informal de correlación entre las áreas estudiadas para los tiempos Oligoceno Tardío-Mioceno Temprano a Medio, basado en presencias de especies características de palinomorfos consideradas como guía. Las asociaciones fueron definidas en base a especies de polen de angiospermas y de dinoquistes, estableciendo un marco general de referencia que, aunque no pretende ser definitivo, ofrece un punto de partida sobre el cual avanzar en futuras investigaciones.
Materiales y Métodos
En los trabajos correspondientes a cada una de las secciones se detallaron los materiales y métodos utilizados. Para la sección correspondiente al tramo del pozo Ra x- 1 que aquí se comunica por primera vez, se usaron los procedimientos estándar para palinología, analizándose solamente la fracción +25 um del residuo obtenido, en la que se concentró el paleomicroplancton.
Los preparados de las secciones comunicadas anteriormente conservarán la sigla CIRGEO Palin, con el número de catálogo correspondiente, aunque a partir de la fecha se encontrarán depositados en la Palinoteca del Museo Bernardino Rivadavia. Los preparados correspondientes a las muestras del pozo Ra x-1 llevarán la sigla SP Dn, y los números de catálogo 1 a 25 y estarán depositados provisoriamente en la colección personal de una de las autoras (SP).
El material analizado por luz transmitida fue fotografiado con película Agfapan 25 y las coordenadas de los ejemplares de polen y esporas ilustrados corresponden al microscopio Leitz Dialux 924193, y las del paleomicroplancton a los microscopios Ortholux 963806(OL) y Orthoplan 939365 (OP).
Polen y esporas
En el entorno del Oligoceno Tardío-Mioceno Medio, en Patagonia comienza a desarrollarse una vegetación relativamente moderna, representada por un espectro esporopolínico de características definidas y que puede diferenciarse fácilmente de las asociaciones más antiguas, paleogenas en general. Se documentan por primera vez representantes de las compuestas (Mutisiapollis viteauensis (Barreda) Barreda), se diversifican las malváceas (Baumannipollis spp., Malvacipolloides spp.), gramíneas (Graminidites spp.), quenopodiáceas (Chenopodipollis chenopodiaceoides (Martin) Truswell) y efedráceas (Equisetosporites claricristatus (Shakmundes) Barreda, E. lusaticus (Krutzsch) Barreda, E. notensis (Cookson) Romero), empezando a evidenciarse los primeros indicios de una incipiente vegetación abierta, herbácea y arbustiva para los tiempos y áreas considerados y que caracterizarán el paisaje del resto del Neogeno. Se reconocen onagráceas (Diporites aspis Pocknall y Mildenhall, Corsinipollenites atlantica Barreda), symplocáceas (Senipites patagonica Barreda) y escalloniáceas (Quintiniapollis estriatulosa Barreda), y también, aunque en forma restringida tanto temporal como arealmente, goodeniáceas (Poluspissusites puntensis Barreda) y palmáceas (Arecipites subverrucatus (Pocknall) Mildenhall y Pocknall). Las áreas boscosas continúan constituyendo una parte importante de la vegetación, con dominio general de podocarpáceas (Podocarpidites spp., Phyllocladidites mawsonii Cookson, Lygistepollenites florinii (Cookson y Pike) Stover y Partridge, entre otras), fagáceas (Nothofagidites spp.) y araucariáceas (Araucariacites australis Cookson). Entre los helechos son características las esporas de pteridáceas (Muricingulisporis chenquensis Barreda), polypodiáceas (Polypodiisporites radiatus Pocknall y Mildenhall, P. inangahuensis (Couper) Potonié emend. Pocknall y Mildenhall), lofosoriáceas (Cyatheacidites annulatus Cookson) y cyatheáceas (Cyathidites spp.); en las cuencas australes también es frecuente Verrucosisporites kopukuensis (Couper) Stover, de afinidad botánica incierta.
A pesar de la clara identidad composicional de las asociaciones esporopolínicas del área para los tiempos considerados en este estudio, se pudieron identificar especies con distribuciones restringidas que permitieron plantear un esquema palinoestratigráfico informal.
Bioestratigrafía y correlación
Los primeros estudios palinológicos efectuados por una de las autoras (VB) se llevaron a cabo en la cuenca del Golfo San Jorge. Para la misma se propuso un esquema informal de biozonación palinológica basado en el espectro esporopolínico. Se reconocieron entonces, para la Formación Chenque, tres palinozonas que abarcaban el intervalo Oligoceno? Superior-Mioceno Medio. Las mismas se denominaron en orden decreciente de edad, Palinozonas A, B, y C respectivamente, con una zona intermedia de transición entre las zonas A y B (Barreda,
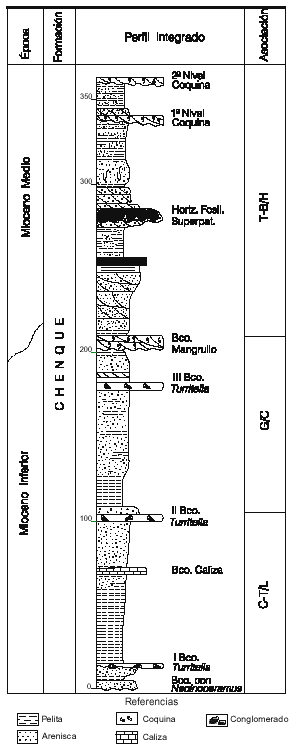
Figura 2. Localidad B, perfil integrado de la Formación Chenque, provincia del Chubut, mostrando la distribución de las asociaciones propuestas /Locality B, composed section of Chenque Formation, Chubut Province, showing the distribution of the proposed associations.
1996). En ese momento no se encontraron argumentos suficientes para descartar una antigüedad de la del Oligoceno para la Palinozona Inferior. Estudios posteriores permitieron reubicar toda la Formación dentro del Mioceno (Palamarczuk y Barreda, 1998). Si bien en un análisis regional más extendido no se mantuvieron constantes todas las distribuciones allí enunciadas, sí se preservó un esquema básico general que permitió una subdivisión del tiempo considerado y el tendido de líneas de correlación para el área. Las mismas fueron utilizadas como base en el esquema que ahora se presenta.
En un primer momento, la edad de las secciones analizadas se estableció teniendo en cuenta, por un lado, las distribuciones de especies características en áreas palinológicamente conocidas y, por otro, las relaciones estratigráficas de las secuencias consideradas. En una etapa posterior, con el análisis de secciones de la cuenca Austral, cuyas edades están más acotadas, ya sea por dataciones radimétricas (Feagle et al., 1995) como por el estudios de otros grupos fósiles, especialmente foraminíferos (Náñez, 1990), se pudo corroborar y en algunos casos ajustar el esquema temporal planteado.
Para el entorno Oligoceno Superior-Mioceno Medio de la costa patagónica y plataforma continental argentina, entre los 45º y 53 º de latitud sur, se definen cuatro asociaciones esporopolínicas, denominadas informalmente: M-M, C-T, G y T-B, y caracterizadas por la presencia de una o más especies indicativas (figura 8). Como marcadores bioestratigráficos se seleccionaron taxones con distribución restringida, poco afectados por controles paleoambientales y/o paleobiogeográficos, y morfologías características que faciliten su identificación.
La asociación M-M, asignada al Oligoceno Tardío, se define por la presencia de las especies de polen Mutisiapollis viteauensis Barreda y Margocolporites tenuireticulatus Barreda y la falta de documentación de taxones registrados en depósitos más modernos. Dentro de esta asociación se producen además, las primeras apariciones, aunque saltuarias, de Baumannipollis variaperturatus Barreda y Rhoipites cienaguensis (Dueñas)Barreda. La documentación constante de estas especies y las apariciones de Cyperaceaepollis neogenicus Krutzsch y Tricolpites trioblatus Mildenhall y Pocknall, asociada a un paulatino aumento en las frecuencias de quenopodiáceas y efedráceas, son utilizadas para definir la base de la asociación C-T a atribuida al Mioceno Temprano. Otras formas de interés bioestratigráfico para la misma, aunque no siempre reconocidas en forma sincrónica son: Corsinipollenites atlantica Barreda y Malvacipolloides comodoroensis Barreda. Los últimos registros de Margocolporites tenuireticulatus y Canthiumidites aff. C. bellus (Partridge) Mildenhall y Pocknall se producen dentro de esta asociación. La primera documentación de Glencopollis ornatus Pocknall y Mildenhall es utilizada para establecer la base de la asociación G, también atribuida al Mioceno Temprano. En un entorno cercano al límite entre las dos asociaciones medias G-T y G las quenopodiáceas y efedráceas alcanzan su punto de acmé. Tubulifloridites antipodica Cookson se registra de manera saltuaria en la parte superior de la asociación G; su presencia continua, junto con los primeros registros de Baumannipollis chubutensis Barreda, se utilizaron para definir la asociación T-B asignada al Mioceno Temprano alto - Mioceno Medio bajo.
Oligoceno Tardío. La asociación M-M, asignada al Oligoceno Tardío, fue identificada en las secciones C y Db (figuras 3 y 5). El espectro polínico recuperado de ambas es comparable aunque no idéntico. En el área sur del Golfo San Jorge (Localidad C) se documentaron familias tropicales que no están presentes en la cuenca Austral (Localidad Db) sugiriendo, ya para el Oligoceno, una diferenciación en fajas climáticas.
Desde el punto de vista bioestratigráfico, en la localidad C, también se documentaron otras formas características, reconocidas en Patagonia sólo en el Oligoceno Superior y Mioceno
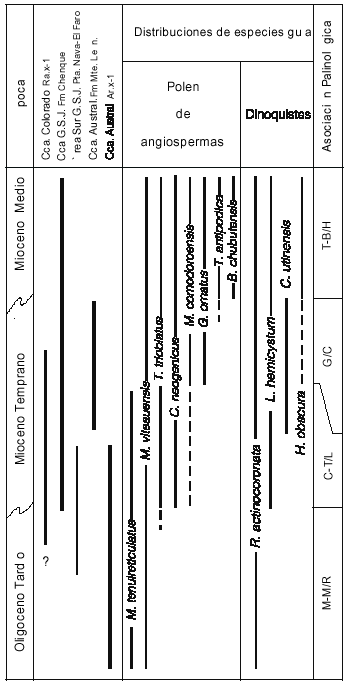
Figura 3. perfil integrado la distribución de las asociaciones propuestas/Locality C, composed section, Punta Nava and El Faro outcrops, Chubut Province, showing the distribution of the proposed associations.
Inferior, como Canthiumidites aff. C. bellus, Diporites aspis, Rubipollis oblatus (Pocknall y Mildenhall) Mildenhall y Pocknall, junto con apariciones saltuarias de Rhoipites cienaguensis y Baumannipollis variaperturatus (Barreda y Palamarczuk, 2000a).
En la localidad Db, el Oligoceno Tardío se identificó en el intervalo 530-350mbbp (Palamarczuk y Barreda, 2000). El contenido esporopolínico del sector es muy pobre en número de especies y ejemplares, y, en consecuencia, además de los tipos característicos (Mutisiapollis viteauensis, Margocolporites tenuireticulatus), casi no se encontraron otros elementos de valor bioestratigráfico. Sólo se reconocieron ejemplares aislados de Assamiapollenites incognitus Pocknall y Mildenhall, especie que presenta un registro continuo recién en el Mioceno.
Mioceno Inferior. Las asociaciones C-T y G, atribuidas al Mioceno Temprano, se identificaron en la mayoría de las secciones y en todas las áreas consideradas (B, C, Da y Db). El espectro esporopolínico presentó variaciones composicionales y de frecuencias con respecto a las asociaciones oligocenas, mostrando un claro avance hacia un paisaje abierto, herbáceo y arbustivo. También se observó un retroceso general de las formas tropicales, especialmente en las localidades más australes: C, Da y Db.
En el perfil cerro Chenque y secciones equivalentes (localidad B), se identificaron las dos asociaciones medias C-T y G (figura 2). La asociación C-T, presente en la mitad inferior del cerro, se caracterizó por las presencias de Tricolpites trioblatus, Baumannipollis variaperturatus y Cyperaceaepollis neogenicus , ésta última recién identificada en un nuevo muestreo lateral del perfil.
En el mismo también se reconoció Assamiapollenites incognitus. En la parte superior de la asociación C-T a se produjeron los últimos registros de Margocolporites tenuireticulatus, Canthiumidites aff C. bellus y Arecipites subverrucatus. Las quenopodiáceas y efedráceas alcanzaron en este sector su punto de acmé. La posterior aparición de Glencopollis ornatus marca la base de la asociación G, desarrollada en la mitad superior del perfil cerro Chenque. Otras especies que también aparecen en el sector son: Poluspissusites puntensis, Corsinipollenites atlantica y Malvacipolloides comodoroensis (Barreda 1996).
En la localidad C sólo fue identificada la asociación C-T en el nivel cuspidal del perfil Punta Nava y en el perfil El Faro (figura 3). Aquí se documentó Cyperaceaepollis neogenicus, ejemplares típicos de la especie Tricolpites trioblatus y registros continuos de Rhoipites cienaguensis y Baumannipollis variaperturatus. También se observó un paulatino incremento en la frecuencias de quenopodiáceas y efedráceas, en relación con lo observado en los términos oligocenos de la cuenca. Otras especies de interés son: Corsinipollenites atlantica y Malvacipolloides comodoroensis, así como Arecipites subverrucatus y Canthiumidites aff. C. bellus, también identificadas en el Mioceno de la cuenca del Golfo San Jorge (localidad B). Las similitudes observadas sugieren una estrecha relación temporal entre el tramo inferior de la Formación Chenque y los niveles aflorantes en el perfil El Faro (Barreda y Palamarczuk, 2000a).
En la Formación Monte León (localidad Da) se reconocieron, aunque parcialmente, las dos asociaciones medias C-T y G (figura 4). El límite entre ambas estaría ubicado en los niveles medio superiores de la Formación Monte León donde se registran por primera vez Glencopollis ornatus y Malvacipolloides comodoroensis. También en este sector se produce un marcado incremento en las frecuencias de quenopodiáceas y efedráceas (Barreda y Palamarczuk, 2000b), mostrando un comportamiento similar al observado en la Formación Chenque. En los términos superiores de esta unidad, asignados a la asociación G, se identificaron ejemplares aislados de Tubulifloridites antipodica.
Por último, en el tramo del pozo Aries (Db) asignado al Mioceno Inferior (350-260mbbp) los tipos polínicos reconocidos sugieren su vinculación con la asociación C-T (figura 5). Las especies estratigráficamente más significativas son: Cyperaceaepollis neogenicus, Mutisiapollis viteauensis, Baumannipollis variaperturatus, Corsinipollenites atlantica y Striasyncolpites laxus Mildenhall y Pocknall.
Esta última, de morfología característica y distribución paleogeográfica y cronoestratigráfica reducida (Mildenhall y Pocknall, 1989), no fue documentada en secciones correlacionables de latitudes más bajas (Palamarczuk y Barreda, 2000). Su afinidad botánica con el género Lyparophyllum de la familia Gentianaceae sugiere un desarrollo en áreas templado frías (Mildenhall y Pocknall, op. cit.). Este hecho estaría determinando su restricción al sector más austral de Patagonia. Otras formas identificadas de valor cronoestratigráfico son: Assamiapollenites incognitus y Margocolporites tenuireticulatus. Sus registros permiten relacionar la sección con la asociación C-T y correlacionarla con los niveles inferiores de la Formación Chenque (localidad B) y cuspidales del área sur del Golfo San Jorge (localidad C).
Mioceno Inferior alto-Mioceno Medio bajo. Depósitos de este entorno temporal sólo fueron identificados en Patagonia en la parte superior de la Formación Chenque (perfiles Antena y Baumann), cuenca del Golfo San Jorge (localidad B), (figura 2). En ellos se observó un buen desarrollo de Tubulifloridites antipodica y la aparición de Baumannipollis chubutensis, elementos que definen la asociación T-B. Estas dos especies también fueron documentadas en el Mioceno Medio de la provincia de San Juan, Formación Chinches (Ottone et al., 1998), a pesar de las marcadas diferencias paleoclimáticas reconocidas para ambas regiones, hecho que estaría avalando sus potencialidades como marcadores bioestratigráficos.
En el Mioceno Inferior alto-Mioceno Medio bajo de Patagonia se evidenció una retracción de las formas xerofíticas (anacardiáceas (Striatricolporites gamerroi Archangelsky), efedráceas, poáceas y en menor proporción quenopodiáceas) acompañadas por un avance de elementos tropicales. Esta asociación sugiere un incremento de temperatura y humedad hasta alcanzar un clima cálido y húmedo, tal como fuera sugerido por el estudio de otros grupos fósiles, principalmente vertebrados (Vucetich, 1994; Tauber, 1997; Kramarz, 1998).
Dinoflagelados
La reseña que se brinda comprende datos integrados de información previamente publicada e inédita. Las observaciones inéditas provienen de las localidades A , Pozo Ra x-1, cuenca del Colorado y B, Fm. Chenque, perfil Cañadón Baumann, cuenca del Golfo San Jorge. Los trabajos correspondientes a estas secciones se encuentran en preparación aunque se presenta aquí una síntesis que fundamenta la inclusión de los mismos en el contexto general de este esquema de correlaciones.
El análisis se centró en el logro de conclusiones de carácter bioestratigráfico, en el estado del conocimiento alcanzado hasta el presente por las autoras, y con el objetivo de establecer posibles correspondencias entre las distintas áreas consideradas. Los aspectos paleobiogeográficos, paleoclimáticos y paleoambientales se mencionan sólo cuando afectan de manera directa la distribución temporal de las especies.
Cuenca del Colorado. La sección disponible para el análisis consistió en un intervalo del pozo Ra x –1, localizado en el sector costa afuera de la plataforma continental argentina, coordenadas aproximadas 40º S y 58º O. Para esta misma cuenca, en su sector marginal, entre otros trabajos previos, pueden citarse: Quattrocchio et al., 1986; Guerstein, 1990a y b; Guerstein y Quattrocchio, 1988; Guerstein y Guler, 2000; Archangelsky, 1996. Las muestras fértiles estu-
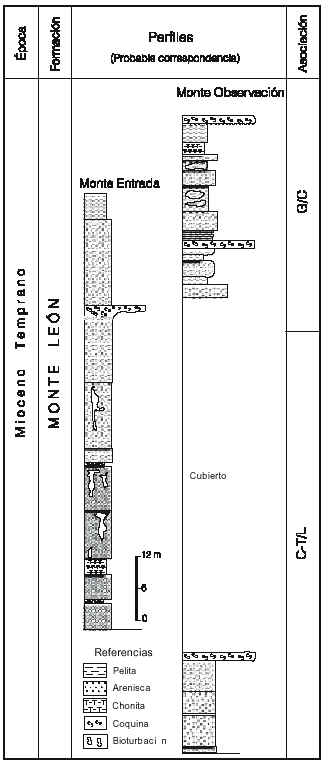
Figura 4. Localidad Da, Formación Monte León, secciones Monte Entrada y Monte Observación, provincia de Santa Cruz indicando la distribución de las asociaciones propuestas/Locality Da, Monte León Formation, Monte Observación and Monte Entrada sections, Santa Cruz Province, showing the distribution of the the proposed associations.
diadas, todas provenientes de tramos de cutting, cubrieron las profundidades 1930 a 1610mbbp (figura 6) abarcando 190m cuspidales de la Formación Elvira, hasta la profundidad de 1740m, donde se da el pase a la Formación Barranca Final (Gamerro y Archangelsky, 1981), que se analizó en sus 130m basales. Por debajo de este intervalo sólo se revisó una muestra correspondiente al nivel 2030m que resultó estéril para palinología. En la profundidad de 1610m se registró un pulso marino extraordinariamente rico en dinoquistes, casi en un 100% de afinidad gonyaulacoide, constituido mayormente por representantes del grupo de los espiniféridos, con una gran variedad morfológica, comprendiendo los géneros Spiniferites, Achomosphaera, Haphniasphaera e Hystrichostrogylon. El segundo grupo dominante identificado fue el del complejo Operculodinium, con presencia de O. centrocarpum (Deflandre y Cookson) Wall, O. giganteum Wall, y un número de variedades de estos tipos básicos. El grupo de especies pequeñas, de características semejantes a O. placitum Drugg y Loeblich, pero también con gran variación morfológica, es un componente importante de la asociación. Otras especies presentes, aunque en un número menor son: Cannosphaeropsis utinensis Wetzel, Melitasphaeridium choanophorum (Deflandre y Cookson) Harland y Hill, Impagidinium spp., Lingulodinium hemicystum Mc Minn, Hystrichokolpoma rigaudiae Deflandre y Cookson, Nematosphaeropsis spp., ?Reticulatosphaera actinocoronata (Benedek) Bujak y Matsuoka, Systematophora placacantha (Deflandre y Cookson) Davey et al., Schematophora sp., Dapsilidinium pseudocolligerum (Stover) Bujak et al., Tectatodinium pellitum Wall y muy escasa presencia de Hystrychosphaeropsis obscura Habib, no documentada en niveles más antiguos del tramo. El contenido en paleomicroplancton disminuye sensiblemente en la muestra inferior, 1630m, registrándose partir de los 1650m nuevamente un aumento del contenido en número de ejemplares, que se mantiene relativamente estable y reflejando condiciones de ambiente de plataforma interna hasta el nivel 1790m, aunque en algunas muestras se observaron leves oscilaciones representativas de condiciones variables de cercanía a la costa. La composición, aunque mucho más pobre que la del nivel 1610 en preservación y número de ejemplares, mantiene una constante que es la dominancia de los grupos asignables a los complejos Spiniferites spp. y Operculodinium spp., además de las presencias, entre otras especies, de Tectatodinium pellitum, Nemathosphaeropsis lemniscata., Melitasphaeridium sp., Hystrichokolpoma spp., Systematophora placacantha, y con participación de formas asignables al grupo de los protoperidiniodes, aunque en mucho menor abundancia que en latitudes más australes. Están presentes los géneros Brigantedinium, Lejeunecysta y Selenopemphix. Hacia la base del tramo fértil se observó la presencia de otras especies del género Lingulodinium, con opérculos complejos, imvolucrando varias paraplacas, aunque en ningún caso pudo asignarse claramente el material a L. machaerophorum (Deflandre y Cookson) Wall. En el nivel 1790m se observaron escasos ejemplares con procesos abiertos distalmemte, comparables a los señalados en Martin, 1991.
No se observó ninguna discontinuidad composicional brusca en los niveles representativos del pase formacional. Algunos registros de importancia bioestratigráfica pudieron documentarse en el tramo considerado. Cannosphaeropsis utinensis no está presente más allá del nivel 1630m, Lingulodinium hemicystum se documentó hasta el nivel 1790 m. Tuberculodinium vancampoae (Rossignol) Wall se registró de manera escasa y muy saltuaria en algunos niveles, ya que es una forma fuertemente controlada por condiciones ambientales y oceanográficas, pero su presencia en la muestra basal, 1930m, fue un criterio utilizado para establecer el límite inferior en la asignación temporal del intervalo palinológicamente fértil, ya que no se lo conoce en depósitos más antiguos que los del Oligoceno Tardío.
Entre los 1770 y los 1790m, se registraron algunos ejemplares del género Deflandrea, en dos tipos básicos, uno comparable a D. phosphoritica Eisenack, y otro de pared del cuerpo central con fuertes granulaciones, mencionado en Biffi y Manum (1988) como D. cf heterophlycta Deflandre y
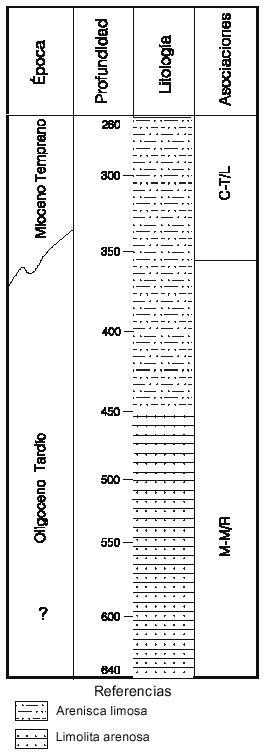
Figura 5. Localidad Db. Representación simplificada del intervalo estudiado del pozo Aries x-1, plataforma continental argentina, mostrando la distribución de las asociaciones propuestas/Locality Db. Simplified representation of the studied interval, Aries x-1 well, Argentine Continental Shelf, showing the distribution of the proposed associations.
Cookson. Esta aparición aquí tiene la doble importancia de ser registrado el género por primera vez en la Formación Elvira, y resultar además un posible criterio de interpretación cronológica de las sedimentitas portadoras. La extinción del género por mucho tiempo se consideró como ocurrida dentro del Oligoceno Tardío, pero trabajos más recientes de detalle en el límite Oligoceno -Mioceno (Brinkhuis et al., 1992) demostraron su presencia con estos tipos morfológicos básicos, en secciones de Italia, país en el que han sido definidos algunos de los estrato tipo del Mioceno, caracterizando el Mioceno Temprano inicial por el acmé de formas afines a las aquí encontradas. Dado que el género no se registró en localidades de Argentina, hasta el presente, en sedimentitas más modernas que las del Oligoceno Temprano, cabe inferir que una presencia moderada en niveles más jóvenes, puede corresponder a una extensión lateral de la zona de acmé en localidades geográficamente alejadas. Lingulodinium hemicystum, reconocido en los mismos niveles, no fue encontrado en depósitos más antiguos que los del Mioceno (Edwards en Head, 1993), lo que apoyaría esta inferencia. En este mismo nivel se observó además un cambio composicional indicativo del entorno del Mioceno Temprano inicial en su transición al Oligoceno. La asociación presentó dominancia de Hystrichokolpoma spp., con presencia de Cordosphaeridium sp. cf C. cantharellus (Brosius) Gocht, Pentadinium laticinctum Gerlach, Saturnodinium sp. y un ejemplar de Sumatradinium sp., comparable al registrado en el pozo Aries x-1 en los dos niveles más modernos estudiados (Palamarczuk y Barreda, 2000). Por debajo del nivel 1790m, las asociaciones se empobrecen marcadamente, por lo que los registros estratigráficos pierden significación. La presencia de Tuberculodinium vancampoae el nivel basal fértil (1930m) acotaría el tramo como no más antiguo que Oligoceno Tardío. El nivel 2100 resultó ya palinológicamente estéril. Se concluye entonces, para el intervalo con influencia marina, una asignación temporal comprendida entre el ?Oligoceno Tardío alto y el Mioceno Temprano alto. Los depósitos suprayacentes no se analizan aquí ya que no se dispone de muestras del sector correspondiente.
Las autoras no han efectuado aún estudios palinógicos de las Formaciones Elvira y Barranca Final en sus áreas tipo. Estudios de microfósiles calcáreos las atribuyen al Oligoceno Inferior y al Entrerriense respectivamente (Malumian et al., 1998a y b). No puede establecerse, por lo tanto, palinológicamente, ninguna relación entre las secciones aquí estudiadas y sus homónimas de las secciones tipo, ya que las edades sugeridas por los palinomorfos no son completamente coincidentes con las propuestas para dichas áreas en los trabajos mencionados. En el caso de la Formación Barranca Final es probable que la diferencia de edades se deba más bien a la falta, en este estudio, de las muestras correspondientes a los términos superiores de la Formación, que posiblemente representen los niveles correlacionables con la sección aflorante en el área tipo.
Perfil Baumann. Se estudió una sección de 64m de la Fm Chenque, expuesta en el Cañadón Baumann. La misma brindó material palinológico continental y marino. El contenido esporopolínico fue comunicado en Barreda (1993, 1996), mientras que la información del paleomicroplancton se comunica por primera vez en esta contribución, sólo en referencia a los tipos de interés bioestratigráfico. Los niveles de mayor contenido en dinoquistes 7B y 8B (figura 7) brindaron asociaciones compuestas mayormente por Hystrichosphaeropsis obscura, ya en su momento de acmé, y Tuberculodiniun vancampoae, con excelente preservación y en gran número de ejemplares, siendo la abundancia en este caso, probablemente, debida a condiciones referibles a una masa de agua de tipo templado cálido. Cabe destacar aquí, además, la presencia de Palaeocystodiniun golzowense Alberti, especie que tiene una muy diferente historia en las cuencas del hemisferio norte que en las de alta latitud del hemisferio sur. En éstas no ha sido documentado en ningún caso en sedimentitas más jóvenes que las del Paleoceno Superior, siendo especie guía
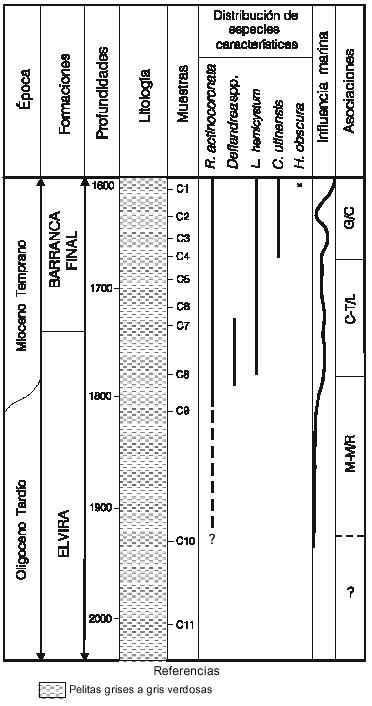
Figura 6. Localidad A. Representación simplificada del intervalo estudiado del pozo Ra x-1, mostrando la distribución de las asociaciones de dinoflagelados propuestas /Locality A. Simplified representation of the studied interval, Ra x-1 well, Argentine Continental Shelf, showing the distribution of the dinoflagellate associations proposed.
para estos tiempos en Nueva Zelanda (Wilson, 1988; Strong et al. 1995). Lo mismo sucede en las distintas secciones de estas edades estudiadas en Tierra del Fuego (Palamarczuk y Olivero, en prensa). La documentación en el Mioceno, con presencia escasa pero significativa, además de un sólo registro en el nivel 1630m de la localidad A, relaciona la sección con corrientes provenientes del norte, en donde P. golzowense es característico, y fue definido, en depósitos del Oligoceno (Alberti, 1961). Se registró además, la presencia abundante de formas referibles en sentido amplio al complejo Systematophora sp. cf S. placacantha, similares a las citadas para el perfil Monte Observación, especialmente en su sector basal, aunque en éste con presencia de algunos ejemplares con unión distal de los complejos de procesos, asignados tentativamente a ?Emmetrocysta sp. (Barreda y Palamarczuk 2000a). Este grupo se diferencia en forma y tamaño de los ejemplares recuperados en la cuenca del Colorado y en el Cerro Chenque, cuyos caracteres bien definidos permiten referirlos a la especie tipo del Mioceno Medio de Australia (Deflandre y Cookson, 1955). En el perfil Cañadón Baumann, la población recuperada consiste en ejemplares algo más pequeños y con gran variabilidad intraespecífica. El valor bioestratigráfico de este complejo deberá ser probado a partir de un mayor número de observaciones. La variabilidad del mismo fue documentada de igual modo para depósitos del Mioceno de Australia estrechamente comparables (Martín, 1991).
Bioestratigrafía y correlación
Considerando el área geográfica entre las latitudes 40º sur y 53º sur, se puede elaborar un esquema preliminar de distribuciones estratigráficas de especies guía, integradas con las asociaciones esporopolínicas (figura 8) caracterizando, a partir de las mismas, cuatro asociaciones informales, y asignándolas tentativamente a entornos cronológicos determinados. Asociación R: identificada en las localidades A? (por debajo de los 1790mbbp.), C (perfil Punta Nava) y Db (por debajo de los 350mbbp). La especie que la caracteriza es ?Reticulatosphaera actinocoronata, y es asignada al Oligoceno Tardío. asociación L, caracterizada por Lingulodinium hemicystum, asignada al Mioceno Temprano y definida en las localidades A (por encima de los 1790mbbp.), B (perfil cerro Chenque parte inferior), C (perfil El Faro), Da (perfiles Monte Entrada y Monte Observación, parte inferior) y Db (350-260 mbbp.). asociación C, Cannosphaeropsis utinensis, del Mioceno temprano alto, localidades A (a partir de los 1630mbbp), B (perfil Cerro Chenque) y Da (perfiles Monte Entrada y Monte Observación, parte superior), y asociación H, con acmé de Hystrichosphaeropsis obscura, en el entorno Mioceno Inferior alto-Mioceno Medio, localidad B (perfil Baumann).
El paleomicroplancton de las edades analizadas es muy rico y potencialmente apto para el establecimiento de una zonación fina, basada en un número suficiente de observaciones y una metolología adecuada. Entre las especies que con toda seguridad serán buenas indicadoras de zona se cuentan entre otras, Nematosphaeropsis rigida Wrenn, Spiniferites mirabilis (Rossignol) Sarjeant, Brigantedidium spp. y Selenopemphix nephroides Benedek, Hystrichostrogylon sp., y Sumatradinium sp. Para estas y otras potenciales especies guías no fue posible todavía determinar con claridad el grado de control ambiental y oceanográfico de sus distribuciones.
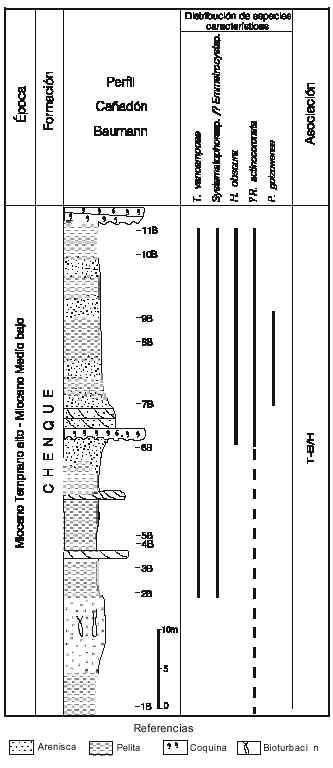
Figura 7. Localidad B, Formación Chenque, perfil Cañadón Baumann mostrando especies seleccionadas de dinoquistes y la distribución de las asociaciones propuestas /Locality B, Chenque Formation, Cañadón Baumann Section, Chubut Province, showing selected dinocysts species and the distribution of the proposed dinoflagellate associations.
Resultados integrados
Los estudios independientes de ambos grupos de palinomorfos han probado que éstos son buenos indicadores bioestratigráficos y por lo tanto, elementos potencialmente útiles en el establecimiento de correlaciones. Además, según se desprende del análisis de depósitos de los tiempos aquí considerados, las distribuciones de especies tomadas como guía son aproximadamente coincidentes para los tipos polínicos y para los dinoquistes lo que permitió proponer un esquema integrado de asociaciones para ambos grupos. Así pudo establecerse la correspondencia de la Asociación M-M/R definida por las presencias conjuntas de Mutisiapollis viteauensis, Margocolporites tenuireticulatus y ?Reticulatosphaera actinocoronata, la asociación C-T/L, con Cyperaceaepollis neogenicus, Tricopites trioblatus y Lingulodinium hemicystum, la asociación G/C, con Glencopollis ornatus y Cannosphaeropsis utinensis, y la asociación T-B/H, con Tubulifloridites antipodica, Baumannipollis chubutensis y acmé de Hystrichosphaeropsis obscura (figura 8).
Conclusiones
La información disponible a partir de distintas disciplinas para los tiempos y localidades aquí analizados es amplia y en muchos casos contradictoria. Las conclusiones a las que se arriba en este trabajo son el resultado de observaciones e interpretaciones personales de las autoras, ya que un tratamiento exhaustivo de la bibliografía previa sobre el tema excedería ampliamente los objetivos planteados en esta contribución.
La palinología es una rama de la paleontología de aplicación relativamente reciente en bioestratigrafia, por lo que es aún mucho lo que resta investigar, ya sea en número de secciones a analizar como en la uniformidad de criterios, especialmente, los taxonómicos, para el tratamiento de las especies. Es por este motivo que las conclusiones que se expresarán a continuación representan sólo un esquema preliminar de correlación que con toda seguridad podrá ser sustancialmente mejorado a partir de observaciones posteriores.
• En base a análisis integrado de los palinomorfos continentales y marinos, ya sea por sus presencias o por sus abundancias relativas, se postula para las latitudes analizadas desde los 53º S hacia el norte, hasta los 40º S, el establecimiento de una transgresión marina que comenzaría en el entorno del Oligoceno Tardío, con manifestaciones de distinta intensidad, y con una extensión estratigráfica variable según las condiciones geológicas de las cuencas en las que se registra.
Esta transgresión se intensificaría a partir del Mioceno Temprano, con distintas expresiones ambientales, que comprenden depósitos costaneros, de plataforma interna, externa, hasta condiciones oceánicas. Luego de un periodo de retracción del nivel del mar, un nuevo pulso, probablemente en el Mioceno Medio, se registra en algunas de las localidades estudiadas.
• Se proponen cuatro asociaciones palinológicas informales, basadas en presencias características de granos de polen de angiospermas y quistes de dinoflagelados: - asociación M-M/R, asignada al OligocenoTardío, y caracterizada por las especies Mutisiapollis viteauensis, Margocolporites tenuireticulatus y ?Reticulatosphaera actinocoronata.
- asociación C-T/L y asociación G/C, del Mioceno Temprano, con presencia de Cyperaceaepollis neogenicus, Tricolpites trioblatus y Lingulodinium hemicystum en la primera, y de Glencopollis ornatus y Cannosphaeropsis utinensis en la segunda.
- asociación T-B/H, con Tubulifloridites antipodica, Baumannipollis chubutensis y acmé de Hystrichosphaeropsis obscura, probablememte ya del Mioceno Medio.
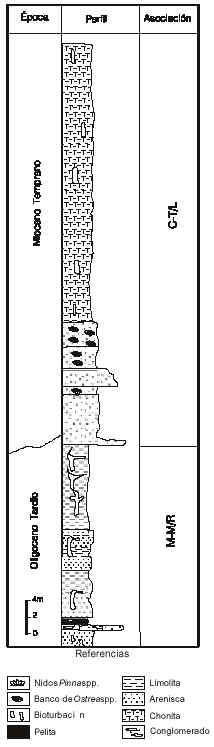
Figura 8. Representación esquemática de correlación entre las áreas estudiadas, mostrando las especies guía de polen de angiospermas y dinoquistes y las asociaciones palinológicas integradas, definidas en este trabajo /Schematic representation of correlation between the studied localities, showing key species of angiosperm pollen and dinocysts and the integrated palynological associations defined in this study.
• Para ambos dominios, continental y marino, se registran diferencias en fajas latitudinales climáticas y de condiciones de aguas, con oscilaciones temporales para cada uno de los momentos considerados.
Agradecimientos
Se agradece especialmente al Dr. Juan Carlos Gamerro por su apoyo constante y por la toma de algunas fotomicrografías de paleomicroplancton. Al Dr. Juan Tramezzani por las facilidades brindadas para la realización de este trabajo, al Dr. L. de Verteuil por la comparación de ejemplares de Brigantedinium de las secciones del Sur del Golfo San Jorge y del pozo Aries x-1, a la agrimensora Amalia González por la confección de las ilustraciones, al Sr. Angel Fusaro por las copias fotográficas y a la Sra. Shelley Hay por la revisión del abstract. El trabajo es una contribución al Proyecto Cenozoico de la plataforma continental argentina y Patagonia (PICT 07-00000-01809. BID 802/OC-AR).
Figura 9.
A, Chenopodipollis chenopodiaceoides (Martin) Truswell. Asociaciones M-M/R, C-T/L, G/C y T-B/H. Oligoceno Tardío-Mioceno Medio; ejemplar de Localidad B, CIRGEO Palin 849: 35,3/95,7/M-M-/R, C-T/L, G/C y T-B/H Associations. Late Oligocene-Middle Miocene; specimen from Locality B.
B, Canthiumidites aff. C. bellus (Partridge) Mildenhall y Pocknall. Asociaciones M-M/R y C-T/L. Oligoceno Tardío- Mioceno Temprano; ejemplar de Localidad B, CIRGEO Palin 843: 44,8/98,3/M-M/R and C-T/L Associations. Late Oligocene-Early Miocene; specimen from Locality B.
C-D, Margocolporites tenuireticulatus Barreda. Asociaciones Inferior y Media a. Oligoceno Tardío-Mioceno Temprano; C, ejemplar de Localidad B, CIRGEO Palin 843: 46,9/106,8; D, ejemplar de Localida Db, CIRGEO Palin 1245: 27/102,8/M-M/ R and C-T/L Associations. Late Oligocene-Early Miocene; C, specimen from Locality B; D, specimen from Locality Db.
E, Malvacipolloides comodoroensis Barreda. Asociaciones C-T/L?, G/C, y T-B/H. Mioceno Temprano a Medio; ejemplar de Localidad B, CIRGEO Palin 957: 31,4/106,6/Middle a?, Middle b and Upper Associations. Early through Middle Miocene; specimen from Locality B.
F, Diporites aspis Pocknall y Mildenhall. Asociaciones M-M/R y C-T/L. Oligoceno Tardío-Mioceno temprano; ejemplar de Localidad B, CIRGEO Palin 1245: 39,4/111,5/M-M/R and C-T/L Associations. Late Oligocene-Early Miocene; specimen from Locality B.
G, Baumannipollis variaperturatus Barreda. Asociaciones M-M/R, C-T/L, G/C y T-B/H. Oligoceno Tardío- Mioceno Medio; ejemplar de Localidad B, CIRGEO Palin 921: 29,3/106/M-M/R, C-T/L, G-C and T-B/H Associations. Late Oligocene-Middle Miocene; specimen from Locality B.
H-I, Equisetosporites claricristatus (Shakmundes) Barreda. Asociaciones M-M/R, C-T/L, G/C y T-B/H. Oligoceno Tardío-Mioceno Medio; ejemplar de Localidad B, CIRGEO Palin 845: 47,2/103,5/ M-M/R, C-T/L, G/C and T-B/H Associations. Late Oligocene-Middle Miocene; specimen from Locality B.
J, Poluspissusites puntensis Barreda. Asociaciones C-T/L? y G/C. Mioceno Temprano; ejemplar de Localidad B, CIRGEO Palin 857: 36,7/109,8/C-T/L? and G/C Associations. Early Miocene; specimen from Locality B.
K, Polypodiisporites radiatus Pocknall y Mildenhall. Asociaciones M-M/R, C-T/L, G/C y T-B/H. Oligoceno Tardío-Mioceno Medio; ejemplar de Localidad B, CIRGEO Palin 898: 28,3/94,8/ M-M/R, C-T/L, G/C and T-B/H Associations. Late Oligocene-Middle Miocene; specimen from Locality B.
L, Arecipites subverrucatus (Pocknall) Mildenhall y Pocknall. Asociaciones M-M/R y C-T/L . Oligoceno Tardío-Mioceno Temprano; ejemplar de Localidad B, CIRGEO Palin 847: 42,8/104,7/ M-M/R and C-T/L Associations. Late Oligocene-Early Miocene; specimen from Locality B.
M, Mutisiapollis viteauensis (Barreda)Barreda. Asociaciones M-M/R, C-T/L, G/C y T-B/H. Oligoceno Tardío- Mioceno Medio; ejemplar de Localidad B, CIRGEO Palin 844: 46,6/96,4/ M-M/R, C-T/L, G/C and T-B/H Associations. Late Oligocene-Middle Miocene; specimen from Locality B.
Todas las figuras x 1260/All figures x 1260.
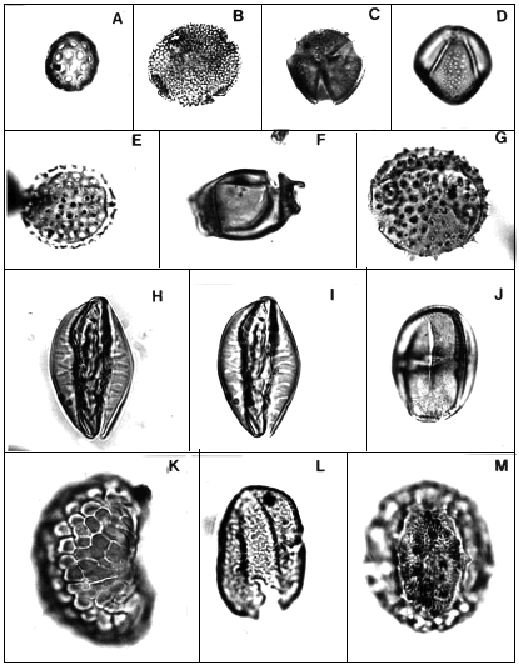
Figura 10.
A- B, Tubulifloridites antipodica Cookson. Asociación T-B/H. Mioceno Temprano alto-Mioceno Medio; (ejemplares de Localidad B); A, CIRGEO Palin 921: 28,6/95,1; B, CIRGEO Palin 921: 41,7/104/T-B/H Association. Late Early-Middle Miocene; specimens from Locality B.
C-D, Striasyncolpites laxus Mildenhall y Pocknall. Asociación C-T/L. Mioceno Temprano; ejemplar de Localidad Db, CIRGEO Palin 1245: 27,2/108/C-T/L Association. Early Miocene; specimen from Locality Db.
E-F, Tricolpites trioblatus Mildenhall y Pocknall. Asociaciones C-T/L, G/C y T-B/H. Mioceno Temprano a Medio; (ejemplares de Localidad B);, E, CIRGEO Palin 921: 27,5/96,2; F, CIRGEO Palin 911: 49/106,9/C-T/L, G/C and T-B/HAssociations. Early through Middle Miocene; specimen from Locality B.
G-H, Glencopollis ornatus Pocknall y Mildenhall. Asociación G/C y T-B/H. Mioceno Temprano a Medio; (ejemplares de Localidad B); G, CIRGEO Palin 882: 48,8/109,8; H, CIRGEO Palin 947: 39,6/107/G/C and T-B/H Associations. Early through Middle Miocene; specimens from Locality B.
I-J, Baumannipollis chubutensis Barreda. Asociación T-B/H. Mioceno Temprano alto-Mioceno Medio; (ejemplares de Localidad B); I, CIRGEO Palin 958: 40,9/104,8; J, CIRGEO Palin 947: 25,5/105/T-B/H Association. Late Early - Middle Miocene; specimens from Locality B.
K-L, Cyperaceaepollis neogenicus Krutzsch. Asociaciones C-T/L, G/C y T-B/H. Mioceno Temprano a Medio; K, ejemplar de Localidad Da, CIRGEO Palin 1218: 55,5/107,8; L, ejemplar de Localidad Db, CIRGEO Palin 1243: 46/97/C-T/L, G/C and T-B/H Associations. Early through Middle Miocene; K, specimen from Locality Da; L, specimen from Locality Db.
M, Corsinipollenites atlantica Barreda. Asociaciones M-M/R, C-T/L, G/C y T-B/H. Oligoceno tardio alto-Mioceno Medio; ejemplar de Localidad B, CIRGEO Palin 958: 46,7/94,4/M-M/R, C-T/L, G/C and T-B/H Associations. Later Oligocene-Middle Miocene; specimen from Locality B.
Figuras A-G y K-L, x 1260; figuras I-J, x 630/Figures A-G and K-L, x 1260; figures I-J, x 630.
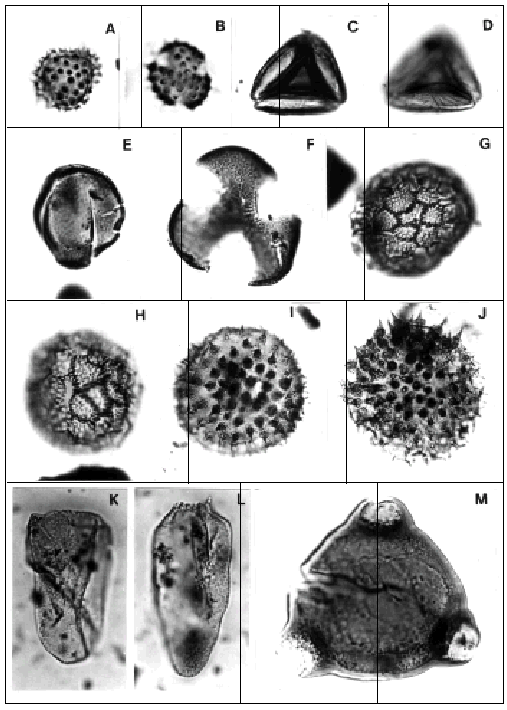
Figura 11
A, Rhoipites cienaguensis (Dueñas) Barreda. Asociaciones M-M/R, C-T/L, G/C y T-B/H. Oligoceno Tardío-Mioceno Medio; fotomicrografía al MEB, (ejemplar de Localidad B)/M-M/R, C-T/L, G/C and T-B/H Associations. Late Oligocene-Middle Miocene; SEM photomicrograph, (specimen from Locality B).
B, Mutisiapollis viteauensis (Barreda) Barreda. Asociaciones M-M/R, C-T/L, G/C y T-B/H. Oligoceno Tardío-Mioceno Medio; fotomicrografía al MEB, (ejemplar de Localidad B)/M-M/R, C-T/L, G/C and T-B/H Associations. Late Oligocene-Middle Miocene; SEM photomicrograph, (specimen from Locality B).
C, F, Tubulifloridites antipodica Cookson. Asociación T-B/H. Mioceno Temprano alto-Mioceno Medio; fotomicrografías al MEB, (ejemplares de Localidad B)/T-B/H Association. Late Early-Middle Miocene; SEM photomicrographs, (specimens from Locality B).
D, Malvacipolloides comodoroensis Barreda. Asociación C-T/L?, G/C y T-B/H. Mioceno Temprano a Medio; fotomicrografía al MEB, (ejemplar de Localidad B)/C-T/L?, G/C and T-B/H Associations, Early through Middle Miocene; SEM photomicrograph, (specimen form Locality B).
E, Glencopollis ornatus Pocknall y Mildenhall. Asociaciones G/C y T-B/H. Mioceno Temprano a Medio; fotomicrografía al MEB, (ejemplar de Localidad B)/G/C and T-B/H Associations, Early through Middle Miocene; SEM photomicrograph, (specimen from Locality B).
G, Equisetosporites claricristatus (Shakmundes) Barreda. Asociaciones M-M/R, C-T/L, G/C y T-B/H. Oligoceno Tardío-Mioceno Medio; fotomicrografía al MEB, (ejemplar de Localidad B)/M-M/R, C-T/L, G/C and T-B/H Associations. Late Oligocene-Middle Miocene; SEM photomicrograph, (specimen from Locality B).
H, Chenopodipollis chenopodiaceoides (Martin) Truswell. Asociaciones M-M/R, C-T/L, G/C y T-B/H. Oligoceno Tardío-Mioceno Medio; fotomicrografía al MEB, (ejemplar de Localidad B)/M-M/R, C-T/L, G/C and T-B/H Associations. Late Oligocene-Middle Miocene; SEM photomicrograph, (specimen from Locality B).
I, Tricolpites trioblatus Mildenhall y Pocknall. Asociaciones C-T/L, G/C y T-B/H. Mioceno Temprano a Medio; fotomicrografía al MEB, (ejemplar de Localidad B)/ C-T/L, G/C and T-B/H Associations. Early through Middle Miocene; SEM photomicrograph, (specimen from Locality B).
J, Arecipites subverrucatus (Pocknall) Mildenhall y Pocknall. Asociaciones M-M/R y C-T/L. Oligoceno Tardío-Mioceno Tremprano; fotomicrografía al MEB, (ejemplar de Localidad B)/M-M/R and C-T/L Associations. Late Oligocene-Early Miocene; SEM photomicrograph, (specimen from Locality B).
K, Corsinipollenites atlantica Barreda. Asociaciones M-M/R, C-T/L, G/C y T-B/H. Oligoceno Tardío-Mioceno Medio; fotomicrografía al MEB, (ejemplar de Localidad B)/M-M/R, C-T/L, G/C and T-B/H Associations. Late Oligocene-Middle Miocene; SEM photomicrograph, (specimen from Locality B).
L, Baumannipollis chubutensis Barreda. Asociación T-B/H. Mioceno Temprano alto-Mioceno Medio; fotomicrografía al MEB, (ejemplar de Localidad B)/T-B/H Association. Late Early-Middle Miocene; SEM photomicrograph, (specimen from Locality B).
Figuras A, B, C, D, H, I, x 1800; figura F, x 2700; figuras E, G, x 1100; figuras J, L, x 600; figura K, x 850/Figures A, B, C, D, H, I, x 1800; figure F, x 2700; figures E, G, x 1100; figures J, L, x 600; figure K, x 850.
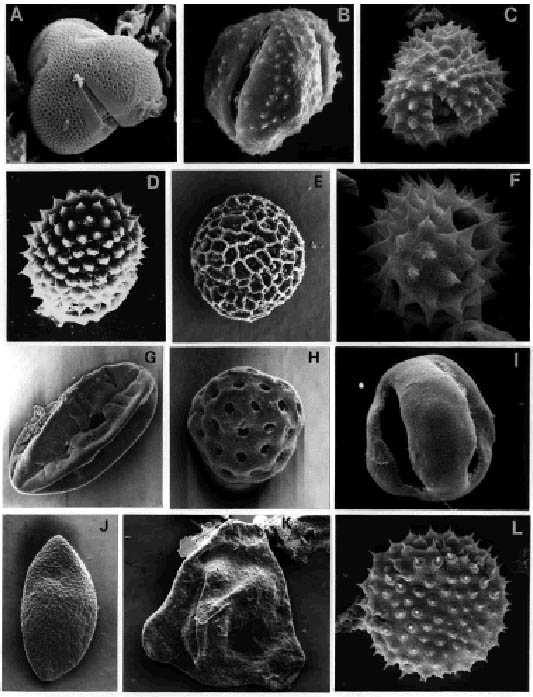
Figura 12
A, Operculodinium giganteum Wall. Asociaciones C-T/L y G/C. Mioceno Temprano; (ejemplar de Localidad A, muestra C1) fotomicrografía al MEB, x 680 /C-T/L and G/C Associations. Early Miocene; (specimen from Locality A, sample C1), SEM photomicrographs, x 680.
B, Melitasphaeridium choanophorum (Deflandre y Cookson) Harland y Hill. Asociación G/C. Mioceno Temprano; (ejemplar de Localidad A, muestra C1); fotomicrografía al MEB, x1500/ G/C Association. Early Miocene; (specimen from Locality A, sample C1); SEM photomicrograph, x 1500.
C, Nematosphaeropsis lemniscata Bujak. Asociaciones C-T/L y G/C. Mioceno Temprano; (ejemplar de Localidad A, muestra C1); fotomicrografía al MEB, x 1360/C-T/L and G/C Associations. Early Miocene; (specimen from locality A, sample C1); SEM photomicrograph, x 1360.
D, Nematosphaeropsis rigida Wrenn. Asociaciones C-T/L y G/C. Mioceno Temprano; (ejemplar de Localidad A, muestra C1); fotomicrografía al MEB, x 1000/C-T/L and G/C Associations. Early Miocene; (specimen from Locality A, sample C1); SEM photomicrograph, x 1000.
E, F, Melitasphaeridium sp. Asociación C-T/L. Mioceno Temprano; (ejemplar de Localidad A, muestra C4); fotomicrografía al MEB, E, x1300; F, detalle de proceso, x 4500/C-T/L Association. Early Miocene; (specimen from Locality A. sample C4); SEM photomicrographs, E, x 1300; F, detail of a process, x 4500.
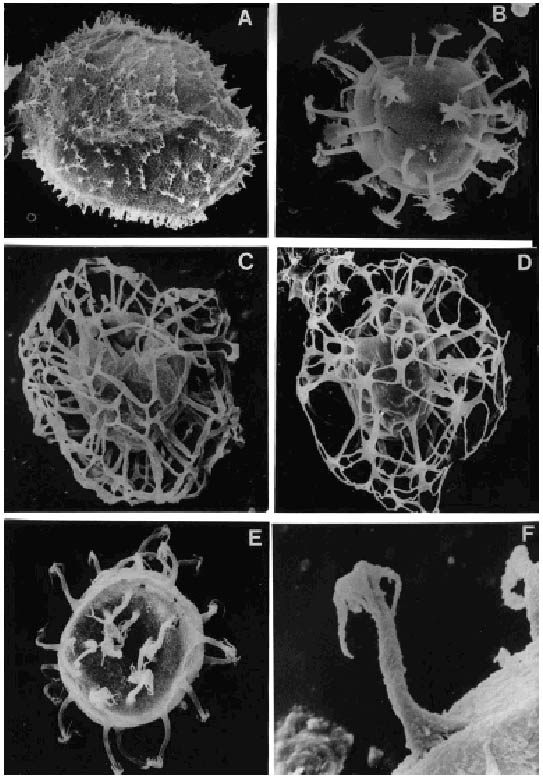
Figura 13
A, D, F, J, Lingulodinium hemicystum Mc Minn. Asociaciones C-T/L y G/C. Mioceno Temprano/C-T/L and G/C Associations. Early Miocene; A, ejemplar mostrando el hemiquiste; (Localidad B), CIRGEO Palin 834: 34,6/107,8 (OL), x 750./ specimen showing the hemicyst; (Locality B); D, F, ejemplar en dos enfoques mostrando el arqueopilo epicistal en enfoque alto y el hemiquiste en enfoque bajo; (Localidad A, muestra C1), Prep.SP Dn 2: 40,4/95,4 (OP), x 740/Specimen showing the hemicystal archeopyle in high focus and the hemicyst in low focus.(Locality A, sample C1); J, Fotomicrografía al MEB; (Localidad B), x 700/ SEM photomicrograph, (Locality B).
B, E, C, Cannosphaeropsis utinensis Wetzel. Asociación G/C. Mioceno Temprano/G/C Association. Early Miocene; B, E, (ejemplares de Localidad A, muestra C1), Prep. SP Dn 3: 52,3/100,4 (OP), x 590/(specimens from Locality A, sample C1); C, fotomicrografía al MEB; (Localidad A, muestra C1), x 980/SEM photomicrograph, (Locality A, sample C1).
G, H, I, ? Reticulatosphaera actinocoronata (Benedek) Bujak y Matsuoka. Asociaciones M-M/R, C-T/L, G/C y T-B/H. Oligoceno Tardío - Mioceno Medio/M-M/R, C-T/L, G/C and T-B/H. Late Oligocene - Middle Miocene; G, foco sobre la expansión distal de los procesos; (ejemplar de Localidad A, muestra C1), Prep. SP Dn 3: 54,9/107,5 (OP), x 750/Focus on process distal expansions; (specimen from locality A, Sample C1); H, (ejemplar de Localidad B, muestra 7B), CIRGEO Palin 952: 52/107,3 (OP), x 750/(specimen from Locality B, sample 7B); I- Fotomicrografía al MEB; (ejemplar de Localidad A, muestra C2), x120./ SEM photomicrograph, (specimen from Locality A, sample C2).
K, L, Hystrichosphaeropsis obscura Habib. Asociación T-B/H. Mioceno Temprano alto-Mioceno Medio/T-B/H Association. Late Early-Middle Miocene; K, enfoque superior dorsal; (ejemplar de Localidad B, muestra 7B), CIRGEO Palin 952: 33/101 (OP) x 740/ High focus on dorsal surface; (specimen from Locality B, sample 7B); L, Mismo ejemplar, corte óptico./Same specimen, optical section.
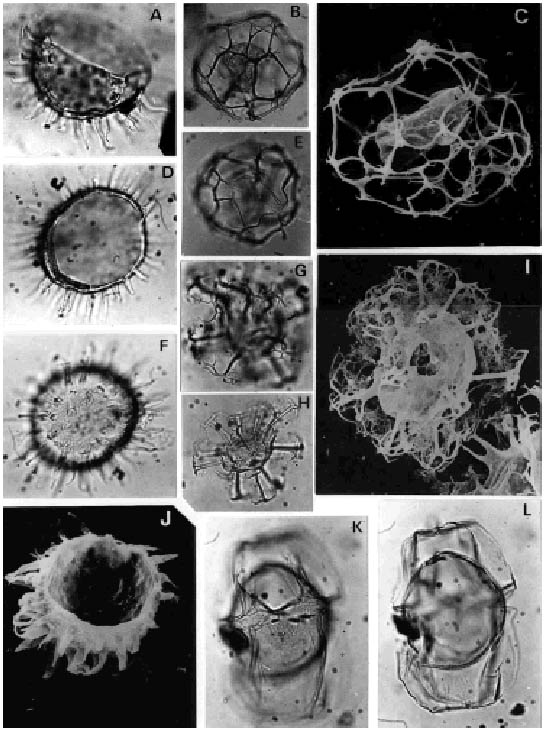
Figura 14
A, Lejeunecysta pulchra Biffi y Grignani. Asociación M-M/R. Oligoceno Tardío (ejemplar de localidad C), CIRGEO Palin 1190: 55,5/97,3 (OL), x 430/M-M/R Association. Late Oligocene; (specimen from Locality C).
B, Deflandrea sp. cf D. phosphoritica Eisenack. Asociación M-M/R. Entorno Oligoceno Tardío-Mioceno Temprano; (ejemplar de Localidad A, Muestra C8), Prep SP Dn 20: 41,9/99 (OP) x 450/M-M/R Association. Late Oligocene –Early Miocene transition. (specimen from Locality A, sample C8).
C, Pentadinium laticinctum Gerlach. Asociación M-M/R. Entorno Oligoceno Tardío-Mioceno Temprano; (ejemplar de Locali-dad A, muestra C8), Prep. SP Dn 20: 48,7/99,6 (OP), x 590/M-M/R Association. Later Oligocene- Early Miocene transition; (specimen from Locality A, sample C8).
D, Sumatradinium sp. Asociación C-T/L. Mioceno Temprano; (ejemplar Localidad Db), CIRGEO Palin 1242: 27,1/94,4 (OL), x 430/C-T/L Association. Early Miocene; (specimen from Locality Db).
E, Deflandrea sp, cf D. heterophlycta Deflandre y Cookson. Asociación M-M/R. Entorno Oligoceno Tardío - Mioceno Temprano; (ejemplar de Localidad A, muestra C8), Prep. SP Dn 20: 39,9/99,3 (OP), x 450/Lower Association , Late Oligocene-Early Miocene transition, (specimen from Locality A, sample C8).
I, Brigantedinium sp. Asociaciones M-M/R, C-T/L y G/C. Oligoceno Tardío-Mioceno Temprano; (ejemplar de Localidad C), CIRGEO Palin 1190: 34,6/95,5 (OL), x 430/M-M/R, C-T/L y G/C Associations. Late Oligocene-Early Miocene; (specimen from Locality C).
F, Deflandrea sp. Asociación M-M/R. Entorno Oligoceno Tardío-Mioceno Temprano. (ejemplar de Localidad A, muestra C8), Prep. SP Dn 20: 41,6/108,4 (OP), x 590/M-M/R Association. Later Oligocene – Early Miocene transition; (specimen from Locality A, sample C8).
G, Palaeocystodinium golzowense Alberti. Asociación C-T/L y T-B/H. Mioceno Temprano a Medio; (ejemplar de Localidad B, muestra7B), CIRGEO Palin 952: 50/93,1, x 450/C-T/L and T-B/H Associations. Early and Middle Miocene; (specimen from locality B, sample 7B).
H, Selenopemphix nephroides Benedek. Asociación C-T/L. Mioceno Temprano; (ejemplar de Localidad Db), CIRGEO Palin 1242: 40,7/102,7 (OL), x 430/ C-T/L Association. Early Miocene; (specimen from Locality Db).
J, Systematophora placacantha (Deflandre y Cookson) Davey et al . Asociaciones C-T/L, G/C y T-B/H. Mioceno Temprano-Medio; (ejemplar de Localidad B, muestra 9B), CIRGEO Palin 954: 29,3/103,2 (OP), x 740/C-T/L, G/C and T-B/H Associations. Early through Middle Miocene. (specimen from Locality B, sample 9B).
K, Distatodinium paradoxus (Brosius) Eaton. Asociación C-T/L. Mioceno Temprano. (ejemplar de Localidad A, muestra C5), Prep.SP Dn 12: 29,3/100,2 (OP), x 740/C-T/L Association. Early Miocene; (specimen from Locality A, sample C5).
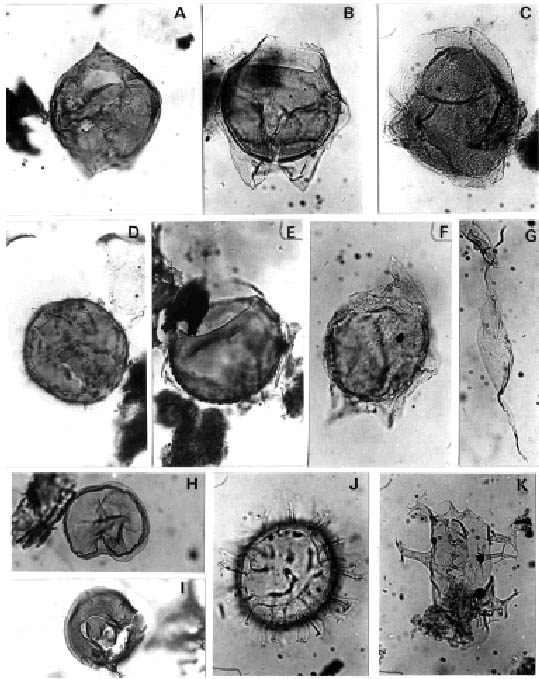
Figura 15
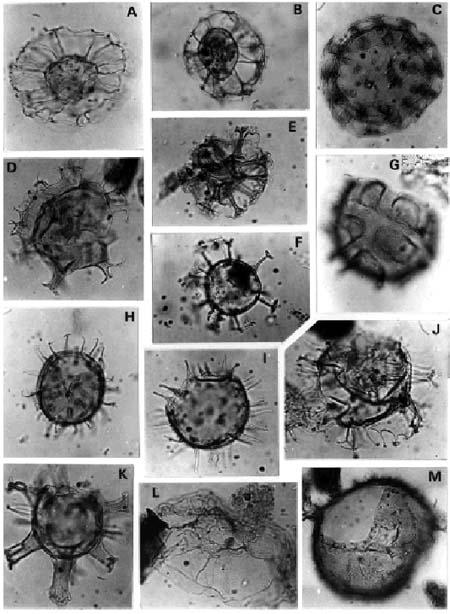
A, Nematosphaeropsis lemniscata Bujak. Asociaciones C-T/L y G/C. Mioceno Temprano. (ejemplar de Localidad A, Muestra C1), Prep. SP Dn 3: 47,8/104,7 (OP), x 740/C-T/L and G/C Associations. Early Miocene (specimen fom Locality A, sample C1).
B, Nematosphaeropsis rigida Wrenn . Asociaciones C-T/L y G/C. Mioceno Temprano. (ejemplar de Localidad Db), CIRGEO Palin 1242: 35,9/105,9 (OL), x 430/ C-T/L and G/C Associations. Early Miocene; (specimen from Locality Db).
C, Tuberculodinium vancampoae (Rossignol) Wall. Asociaciones M-M/R, C-T/L, G/C y T-B/H. Oligoceno Tardío-Mioceno Medio; (ejemplar de localidad B, Muestra. 6B), CIRGEO Palin 947: 27,6/93,2 (OP), x 290/M-M/R, C-T/L, G/C and T-B/ H Associations. Later Oligocene-Middle Miocene; (specimen fom Locality B , sample 6B).
D, Hystrichostrogylon sp. Asociaciones G/C y T-B/H; (ejemplar de Localidad A, muestra C1), Prep.SP Dn 3: 47,6/100,5 (OP), x 590/ G/C and T-B/H Associations. Early through Middle Miocene; (specimen from Locality A, sample C1).
E, Cordosphaeridium sp. cf C. cantharellus (Brosius) Gocht .Asociación M-M/R. Entorno Oligoceno Tardío-Mioceno Tempra-no; (ejemplar de Localidad A, muestra C8), Prep. SP Dn 20: 31,7/98,8 (OP), x 740/M-M/R Association. Later Oligocene – Early Miocene transition; (specimen from Locality A, sample C8).
F, Melitasphaeridium choanophorum (Deflandre y Cookson) Harland y Hill. Asociaciones G/C y T-B/H. Mioceno Temprano a Medio; (ejemplar de Localidad A, muestra C1), Prep.SP Dn 3: 40,5/111 (OP), x 740/G/C and T-B/H Associations. Early through Middle Miocene; (specimen from Locality A, sample C1).
G, Schematophora sp. Asociación C-T/L. Mioceno Tempramo; (ejemplar de Localidad A, muestra C1), Prep. SP Dn 3: 31,9/ 97,7 (OP), x 590/C-T/L Association. Early Miocene; (specimen from Locality A, sample C1).
H, Melitasphaeridium sp. Asociación C-T/L. Mioceno Temprano; (ejemplar de Localidad A, muestra C1), Prep. SP Dn 3: 46,7/ 103,8 (OP), x 740/C-T/L Association. Early Miocene; (specimen from Locality A, sample C1).
I, Dapsilidinium pseudocolligerum (Stover) Bujak et al. Asociaciones C-T/L, G/C y T-B/H. Mioceno Temprano a Medio; (ejemplar de Localidad B, muestra 9B), CIRGEO Palin 954: 31/100,3 (OP), x 740/C-T/L, G/C and T-B/H Associations. Early through Middle Miocene; (specimen from Locality B, sample 9B).
J, Spiniferites mirabilis (Rossignol) Sarjeant . Asociaciones C-T/L, G/C y T-B/H. Mioceno Temprano a Medio; (ejemplar de Localidad B, muestra 10B), CIRGEO Palin 956: 45,5/94,2 (OP), x 740/C-T/L, G/C and T-B/H Associations. Early through Middle Miocene; (specimen from Locality B, sample 10B).
K, Hystrichokolpoma rigaudiae Deflandre y Cookson. Asociaciones C-T/L y G/C. Mioceno Temprano; (ejemplar de Localidad A, muestra C 5), Prep.SP Dn 12: 29,6/ 103,6 (OP), x590/C-T/L and G/C Associations. Early Miocene; (specimen from Locality A, sample C5).
L, Saturnodinium sp. Asociación C-T/L. Entorno Oligoceno Tardío-Mioceno Temprano; (ejemplar de Localidad A, muestra C8), Prep. SP Dn 20: 46,5/108,8 (OP), x 590/C-T/L Association. Later Oligocene- Early Miocene transition; (specimen from Locality A. sample C8).
M, Operculodinium giganteum Wall . Asociación G/C. Mioceno Temprano; (ejemplar de Localidad A, muestra C1), Prep SP Dn 2: 44,7/ 99,3 (OP), x 370/G/C Association. Early Miocene; (specimen from Locality A, sample C1).
Bibliografía
Alberti, G. 1961. Zur Kenntnis mesozoischer und alttertiärer Dinoflagellaten und Hystrichosphaerideen von Nordund Mitteldeutschland sowie einigen anderen europäischen Gebieten. Palaeontographica 116: 1-58.
Archangelsky, S., 1996. Palinoestratigrafía de la plataforma continental. En: Ramos, V. A. y Turic, M.A. (eds.). Geología y Recursos Naturales de la Plataforma Continental Argentina. 13º Congreso Geológico argentino y 3º Congreso de Exploración de Hidrocarburos. (Buenos Aires), Relatorio 4: 67-72.
Barreda, V.D. 1993. Late Oligocene?-Miocene pollen of the families Compositae, Malvaceae and Polygonaceae from the Chenque Formation, Golfo San Jorge Basin, southeastern Argentina. Palynology 17: 169-186.
Barreda, V.D. 1996. Bioestratigrafía de polen y esporas de la Formación Chenque, Oligoceno tardío?-Mioceno de las provincias de Chubut y Santa Cruz, Patagonia, Argentina. Ameghiniana 33: 35-56.
Barreda, V.D. y Palamarczuk, S. 2000a. Palinoestratigrafía del Oligoceno tardío-Mioceno, en el área sur del Golfo San Jorge, provincia de Santa Cruz, Argentina. Ameghiniana 37: 103-117.
Barreda, V.D. y Palamarczuk, S. 2000b. Palinomorfos continentales y marinos de la Formación Monte León en su área tipo, provincia de Santa Cruz, Argentina. Ameghiniana 37: 3-12.
Becker, D. 1964. Micropaleontología del Superpatagoniense de las localidades Las Cuevas y Monte Entrance (provincia de Santa Cruz). Ameghiniana 3: 319-351.
Becker, D. y Bertels, A. 1980. Micropaleontología de la secuencia terciárica de la perforación Puelche (margen continental argentino). 2º Congreso Argentino de Paleontología y Bioestratigrafía y 1º Congreso Latinoamericano de Paleontología (Buenos Aires, 1978), Actas 2: 315-333.
Bellosi, E. 1990. Formación Chenque: registro de la transgresión patagoniana en la cuenca San Jorge. 11º Congreso Geológico Argentino (San Juan), Actas 2: 57-60.
Bellosi, E. 1995. Paleogeografía y cambios ambientales de la Patagonia central durante el Terciario medio. Boletín de Informaciones Petroleras 44: 50-83.
Bertels, A. 1970. Sobre el "Piso Patagoniano" y la representación de la época del Oligoceno en Patagonia Austral. República Argentina. Revista de la Asociación Geológica Argentina 25: 495-501.
Bertels, A. 1975. Bioestratigrafía del Paleógeno en la República Argentina. Revista Española de Micropaleontología 7: 426-450.
Bertels, A. 1978. Estratigrafía y foraminíferos (Protozoa) bentónicos de la Formación Monte León (Oligoceno) en su área tipo, provincia de Santa Cruz, República Argentina. 2º Congreso Argentino de Paleontología y Bioestratigrafía y 1º Congreso Latinoamericano de Paleontología (Buenos Aires), Actas 2: 213-273.
Biffi, U. y Manum, S.B. 1988. Late Eocene-Early Miocene dinoflagellate cyst stratigraphy from the Marche Region (Central Italy). Bollettino della Società Paleontologica Italiana 27: 163-212.
Brinkhuis, H., Powell, J. y Levenboom, D. 1992. High resolution dinoflagellate cyst stratigraphy of the Oligocene-Miocene transition interval in Northwest and central Italy. En: M. Head and J.H. Wreen (eds.), Neogene and Quaternary dinoflagellate cysts and acritarchs, Dallas, American Association of Stratigraphic Palynologists Foundation, pp. 219-258.
Camacho, H.H. 1974. Bioestratigrafía de las formaciones marinas del Eoceno y Oligoceno de la Patagonia. Anales de la Academia Nacional de Ciencias Exactas, Físicas y Naturales 26: 39-57.
Camacho, H.H. y Fernández, J.A. 1956. La transgresión patagoniense en la costa atlántica entre Comodoro Rivadavia y el curso inferior del río Chubut. Revista de la Asociación Geológica Argentina 9: 23-45.
Cione, A y Expósito E. 1978. Condrichthyes (Pisces) del "Patagoniano" s.l. de Astra, Golfo de San Jorge, provincia de Chubut, Argentina. Su significación paleoclimática y paleobiogeográfica. 2º Congreso Argentino de Paleontología y Bioestratigrafía y 1º Congreso Latinoamericano de Paleontología (Buenos Aires), Actas 2: 275-290.
Deflandre G. y Cookson, I. 1955. Fossil Microplanktom from Australian Late Mesozoic and Tertiary sediments. Australian Journal of freshwater Research 6: 242-313.
Feagle, J.G., Bown, T.M., Swisher, C. y Buckley, G. 1995. Age of the Pinturas and Santa Cruz Formation. 6º Congreso Argentino de Paleontología y Bioestratigrafía, (Trelew), Actas: 129-135.
Feruglio, E. 1949. Descripción geológica de la Patagonia, 2: 1- 545.
Frenguelli, J. 1929. Descripción de algunos perfiles de la zona petrolífera de Comodoro Rivadavia. Boletín de Informaciones Petroleras 59: 575-605.
Gamerro, J.C. y Archangelsky, S. 1981. Palinozonas Neocretácicas y Terciarias de la plataforma continental argentina en la Cuenca del Colorado. Revista Española de Micropaleontología 13: 119-140.
Guerstein, G.R. 1990a. Palinología estratigráfica del Terciario de la cuenca del Colorado. República Argentina. Parte II: especies marinas de la perforación Nadir Nº 1. Revista Española de Micropaleontología 22: 167-182.
Guerstein, G.R. 1990b. Palinología estratigráfica del Terciario de la cuenca del Colorado, República Argentina. Parte III: Estudio sistemático y estadístico de la perforación Puerto Belgrano Nº 20. Revista Española de Micropaleontología 22: 459- 480.
Guerstein, G.R. y Guler, M.V. 2000. Bioestratigrafía basada en quistes de dinoflagelados del Eoceno-Mioceno del pozo (YPF) Ombucta x-1, cuenca del Colorado, Argentina. Ameghiniana 37: 81-90.
Guerstein, G.R. y Quattrocchio, M.E. 1988. Palinozonas e interpretación estratigráfica mediante análisis de agrupamiento del Terciario de la Cuenca del Colorado, República Argentina. 2º Jornadas Geológicas Bonaerenses. (Bahía Blanca). Actas 1: 27-35.
Hatcher, J.B. 1897. Geology of Southern Patagonia. American Journal of Science 4: 327-354.
Head, M.J. 1993. A forum on Neogene and Quaternary dinoflagellate cysts. Palynology 17: 201-239.
Ihering, H. von 1902. Historia de las ostras argentinas. Anales del Museo Nacional de Historia Natural, Buenos Aires, 2: 109-125.
Kramarz, A. 1998. La fauna de roedores de la Formación Pinturas, Mioceno Medio inferior de la provincia de Santa Cruz. 7º Congreso Argentino de Paleontología y Bioestratigrafía (Bahía Blanca). Resúmenes: 67.
Legarreta, L. y Uliana, M.A. 1994. Asociaciones de fósiles y hiatos en el Supracretácico-Neógeno de Patagonia: Una perspectiva estratigráfico-secuencial. Ameghiniana 31: 257-282.
Malumián, N. 1970. Bioestratigrafía del Terciario marino del subsuelo de la provincia de Buenos Aires. Ameghiniana 7: 173- 204.
Malumián, N. 1972. Foraminíferos del Oligoceno y Mioceno del subsuelo de la provincia de Buenos Aires. Ameghiniana 9: 97- 137.
Malumián, N. y Náñez, C. 1988. Asociaciones de foraminíferos del Terciario medio de la cuenca Austral: sus relaciones con eventos eustáticos globales. Revista de la Asociación Geológica Argentina 43: 257-264.
Malumián, N. y Náñez, C. 1991. Paleogeografía del Terciario medio del cono sur: avance de aguas antárticas. 6º Congreso Geológico Chileno (Santiago), Resúmenes expandidos: 847-851.
Malumián, N. y Náñez, C. 1998. El género Transversigerina y la edad de la transgresión patagoniana. 10º Congreso Latinoamericano de Geología y 6º Congreso Nacional de Geología Económica (Buenos Aires), Actas 1: 285-290.
Malumián, N., Náñez, C. y Jannou, G. 1998. La Formación Elvira en su localidad tipo. Cuenca del Colorado. Foraminíferos y edad. 10º Congreso Latinoamericano de Geología y 6º Congreso Nacional de Geología Económica (Buenos Aires) Actas 1: 114-119.
Malumián, N., Suriano, J.M. y Cobos, J.C. 1998b. La Formación Barranca Final en su localidad tipo, Mioceno, cuenca del Colorado. 10º Congreso Latinoamericano de Geología y 6º Congreso Nacional de Geología Económica (Buenos Aires), Actas 1: 125- 130.
Martin, H.A. 1991. Dinoflagellate and spore pollen biostratigraphy of the S.A.D.M.E. MC63 bore, western Murray Basin. Alcheringa 15: 107-144.
Mildenhall, D.C. y Pocknall, D.T. 1989. Miocene-Pleistocene spores and pollen from Central Otago, South Island, New Zealand. New Zealand Geological Survey, Paleontological Bulletin 59: 1-128.
Náñez, C. 1990. Foraminíferos y bioestratigrafía del Terciario medio de Santa Cruz oriental. Revista de la Asociación Geológica Argentina 43: 493-517.
Ortmann, A. 1900. Sinopsis of the collections of invertebrate fossils made by the Princeton Expedition to Southern Patagonia. American Journal of Science. 4: 368-381.
Ottone, E.G., Barreda, V.D. y Pérez, D.J. 1998. Basin evolution as reflected by Miocene palymomorphs from the Chinches Formation, Frontal Cordillera (32º S), San Juan Province, Argentina. Revista Española de Micropaleontología 30: 35-47.
Palamarczuk, S. y Barreda, V. 1998. Bioestratigrafía en base a quistes de dinoflagelados de la Formación Chenque (Mioceno), provincia del Chubut, Argentina. Ameghiniana 35: 415-426.
Palamarczuk, S. y Barreda, V. 2000. Palinología del Paleogeno tardío-Neogeno temprano, pozo Aries x-1, plataforma continental argentina, Tierra del Fuego. Ameghiniana 37: 221-234
Palamarczuk, S. y Olivero, E. 2000. Asociaciones de dinoflagelados del Cretácico Superior-Paleogeno en el área del Río Bueno, costa atlántica de la isla Grande de Tierra del Fuego. 11º Simposio Argentino de Paleobotánica y Palinología (Tucumán), Resúmenes (en prensa).
Parma, S.G. 1985. Eoscutella Grant y Hertlein (Echinodermata: Clypeasteroida) en el Patagoniano (Terciario Inferior) de la Provincia de Santa Cruz. Revista de la Asociación Geológica Argentina 37: 23-49.
Parma, S.G. 1989. El género Platipygus (Echinoidea: Cassiduloida) en el Terciario de la Provincia de Santa Cruz, República Argentina. Ameghiniana 25: 213-224.
Quattrocchio, M., Guerstein, G.R. y Sbardellati, M.S. 1986. Neogene dinoflagellate cysts from the Colorado Basin, Argentina. En: Wrenn, J.; Duffield, S. y Stein, J.A. (eds.). First Symposium on Neogene dinoflagellate cyst biostratigraphy (New Yor). American Association of Stratigraphic Palynologists, Contributions Series 17: 151-157.
Strong, C. P.,Hollis, C. J. y Wilson, G. 1995. Foraminiferal, radiolarian, and dinoflagellate biostratigraphy of Late Cretaceous to Middle Eocene pelagic sediments (Muzzle Group), Mead Stream, Marlborough, New Zealand. New Zealand Journal of Geology and Geophysics 38: 171-212.
Tauber, A. 1997. Paleoecología de la Formación Santa Cruz (Mioceno Inferior) en el extremo sudoeste de la Patagonia. Ameghiniana 34: 517-529.
Vucetich, M.G. 1994. La fauna de roedores de la Formación Cerro Boleadoras (Mioceno Inferior?) en la provincia de Santa Cruz (Argentina). Acta Geológica Leopoldensia 39: 365-374.
Wichmann, R. 1918. Estudios geológicos e hidrogeológicos en la región comprendida entre la boca del río Negro, San Antonio Oeste y Choele Choel. Ministerio de Agricultura de la Nación, Dirección Nacional de Geología y Minería, Anales, 13: 9- 41.
Wilson, G.J. 1988. Paleocene and Eocene dinoflagellate cysts from Waipawa, Hawkes Bay, New Zealand. New Zealand Geological Survey. Paleontological Bulletin 57: 1-96.
Windhausen, A. 1924. Líneas generales de la constitución geológica de la región situada al oeste del Golfo de San Jorge. Boletín de la Academia Nacional de Ciencias, Córdoba, 27: 1-320.
Recibido: 30 de junio de 2000
Aceptado: 1 de noviembre de 2000