The
Ordovician Brachiopod Faunas of Argentina: Chronology and Biostratigraphic
Succession
Juan L. BENEDETTO
CONICET. Cátedra de Estratigrafía y Geología Histórica, Facultad de Ciencias Exactas, Físicas y Naturales,
Universidad Nacional de Córdoba, Av. Vélez Sarsfield 299, 5000 Córdoba, Argentina. jbenedetto@onenet.com.ar
Abstract.
THE ORDOVICIAN
BRACHIOPOD FAUNAS OF ARGENTINA:
CHRONOLOGY AND BIOSTRATIGRAPHIC SUCCESSION.
This article summarizes brachiopod biostratigraphic data from Ordovician basins
of Argentina. Taxonomic composition of the five brachiopod biozones recognized
by Herrera and Benedetto (1991) in the Argentine Precordillera is updated on the
basis of new information accumulated in the past ten years, especially from
silicified material present in the lower part of the San Juan Formation. The Archaeorthis
Zone is subdivided into three ‘subassemblages’ and the new Ranorthis
niquivilensis Zone is recognized. After the drowning of the carbonate
platform sedimentation becomes dominantly clastic and stratigraphic units
underwent strong lithofacial changes through the Precordillera basin. As a
result, late Llanvirn to late Ashgill brachiopod assemblages are comparatively
more restricted in space and time. In the Arenig volcano-sedimentary sequences
of the Famatina Range three distinct brachiopod assemblages can be recognized
(in ascending order): Suriorthis depressus, Mollesella planiventralis and
Famatinorthis turneri. The brachiopod fauna reported from western Puna is
probably coeval with the Famatinian S. depressus assemblage. The Central
Andean brachiopods are still poorly known. On the basis of available data from
the Cordillera Oriental, a preliminary stratigraphic succession of eight
brachiopod assemblages spanning the early Tremadocian to midlle Arenig interval
is proposed.
Resumen.
LAS FAUNAS
DE BRAQUIÓPODOS ORDOVÍCICOS DE ARGENTINA:
CRONOLOGÍA Y SUCESIÓN
BIOESTRATIGRÁFICA.
Este trabajo resume los datos disponibles sobre biostratigrafía de los braquiópodos
ordovícicos de Argentina. Se actualiza la composición taxonómica de las cinco
biozonas de braquiópodos reconocidas por Herrera y Benedetto (1991) en la
Precordillera Argentina en base a la nueva información obtenida en los últimos
diez años, especialamente la proveniente de los braquiópodos silicificados de
la Formación San Juan. La Zona de Archaeorthis es subdividida en tres
‘sub-asociaciones’ y se reconoce la nueva Zona de Ranorthis
niquivilensis. Después de la inundación de la plataforma carbonática la
sedimentación se tornó dominantemente clástica y las unidades estratigráfiacas
experimentan fuertes cambios litofaciales a través de la Precordillera. Como
resultado, las faunas de braquiópodos del intervalo Llanvirniano tardío-Ashgilliano
tardío están comparativamente más restringidas en el tiempo y el espacio que
las del Ordovícico temprano. En la secuencia volcaniclástica Arengiana de la
Sierra de Famatina se han reconocido tres asociaciones de braquiópodos (en
orden ascendente): Suriorthis depressus, Mollesella planiventralis y
Famatonorthis turneri. La fauna de braquiópodos de la Puna occidental
(Vega Pinato) es relativamente similar y probablemente contemporánea de la
asociación de S. depressus de Famatina. Los braquiópodos de la cuenca
Andina Central son aún poco conocidos. Sobre la base de los datos disponibles
de la Cordillera Oriental se propone en forma preliminar una sucesión de ocho
‘asociaciones’ que abarcan el lapso Tremadociano temprano- Arenigiano medio.
Key
words: Ordovician. Brachiopods. Argentina
Palabras
clave: Ordovícico.
Braquiópodos.
Argentina
Introduction
Among
benthic invertebrates, trilobites have been used, to a greater and lesser
extent, for biostratigraphical purposes, especially since the publication of the
Harrington and Leanza (1957) monograph on Ordovician trilobites in which they
proposed a formal biozonation of Ordovician rocks of Argentina. The first
attempt to use brachiopods for a biostratigraphic zoning was carried out in the
thick and continuous succession of fossiliferous limestones of late Tremadocian
to early Llanvirn age (San Juan Formation) exposed everywhere in the
Precordillera basin. The increasing knowledge on taxonomy and stratigraphic
ranges of the abundant and diverse brachiopods from the San Juan Formation led
to propose a biozonal scheme based on five assemblage biozones, each named for
their dominant components (from lowest to highest): Archaeorthis Zone, Huacoella
Zone, Niquivilia Zone, Monorthis Zone and Ahtiella Zone
(Herrera and Benedetto, 1991). Although variations in taxonomic composition
among successisve ‘assemblages’ may have been environmentally controlled, as
their close correspondence with biofacial and lithofacial changes suggests
(Carrera et al., 1999, 2001), they are not time-recurrent. Environmental
changes through deepening and shallowing sequences affect principally the
proportion of higher-taxa. For example, diversity of plectambonitoids attains
higher values in the open shelf settings prevailing during the Huacoella-Niquivilia
biozones and the Llanvirn Ahtiella biozone. Changes among biozones
involving apparition of new genera and species were produced by a combination of
phenomena such as evolution within a given lineage (e.g. the Nothorthis-Ranorthis
lineage, Benedetto, in press b; Benedetto et al., in press),
immigration of new taxa, and extintion of others.
Since
the brachiopod-based biozones were proposed, much additional taxonomic work has
been published (Benedetto, 1998a, in press b, and references in both papers).
Nevertheless, correlation among sections of the San Juan Formation based on
brachiopod zones has still high confidence, especially in the lower and midlle
part of the unit. Correlation of its upper part (Ahtiella Zone) is
somewhat more problematical because of lithofacial and biofacial changes between
sections located in the northern and southern parts of the basin (Sánchez et
al., 1996).
Most
of new taxonomic information comes from the silicified material from the lower
half of the Archaeorthis Zone, the Niquivilia Zone and the Monorthis
Zone (Benedetto, 2001a; 2002; Benedetto et al., in press). A
preliminary refinement of the biostratigraphic succession within the lower part
of the Archaeorthis Zone was recently made by Benedetto et al. (in
press). On the other hand, the highdiversity brachiopod assemblages from both
the Ahtiella Zone and the succeeding Skenidioides assemblage
(defined herein) are actually under revision, thus a substantial increase in
number of taxa can be expected inasmuch as this study is concluded.
In
the Famatina basin (La Rioja and Catamarca provinces) great advances have been
made recently in elucidating both the taxonomic composition and
paleobiogeographic signature of brachiopod faunas. Of particular importance have
been the extensive collections from the volcanoclastic successions exposed in
the Chaschuil and central Famatina regions. As a result, 20 genera and 24
species have been identified and carefully located in measured stratigraphic
sections (Benedetto, 1994, in press a).
Although
most of Central Andean stratigraphic units contain brachiopods, they are still
poorly known. Since the reports of Kobayashi (1937) and Harrington (1938) on
some early Ordovician brachiopods from northwestern Argentina, only a few
taxonomic works have been published (Benedetto, 1998c, 2002; Benedetto and
Carrasco, in press), most of recently collected material has yet to be studied.
For this reason, the stratigraphic succession of brachiopod assemblages given
below only constitutes a first attempt to organize the available data. Three
different kind of faunal associations are considered: (a) formally defined
biostratigraphic zones, (b) informal biostratigraphic units, named here
‘assemblages’ or ‘sub-assemblages’ (e.g. Destombesium assemblage),
and (c) faunas whose biostratigraphic significance has not been demonstrated,
often including poorly preserved, sparse and/or incompletely known taxa (‘not
zoned’).
Chronology
of brachiopod faunas
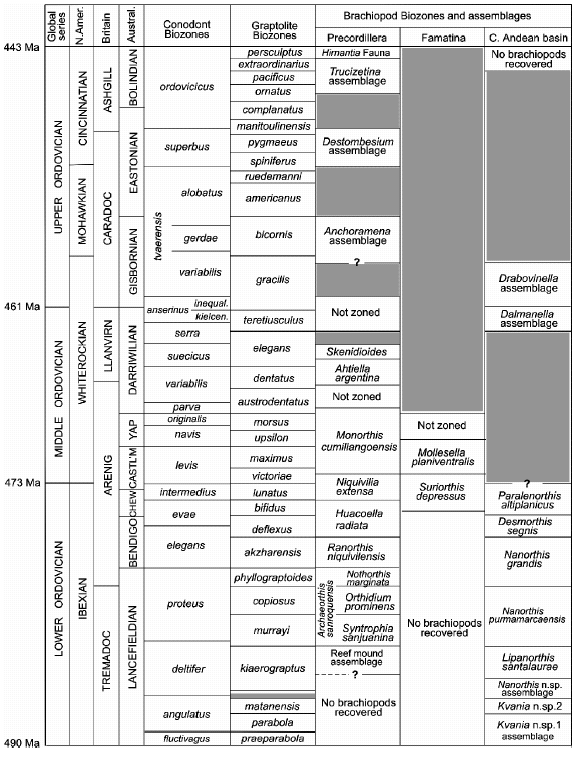
Figure
1 shows the stratigraphic succession of brachiopod biozones and assemblages
recognized in this paper. They have been referred to the standard
chronostratigraphic chart on the basis of conodont and graptolite information.
In the Precordillera basin, conodonts have been essential for dating the San
Juan Formation brachiopod biozones (Lehnert, 1995; Albanesi et al.,
1995b, 1998a, and references therein), while graptolites provide invaluable ties
between carbonate and siliciclastic successions (Albanesi et al., 1999; Ortega
and Albanesi, 1999 and references therein).
In
the Sierra de Famatina, the age of the Suri and Molles formations has been based
largely on conodont and graptolite evidence, and additionally on trilobite
faunas (Vaccari and Waisfeld, 1994).
Graptolites
from the lower member of the Suri Formation described by Toro and Brussa (1997)
belongs to the early-middleArenig B. deflexus and D. bifidus biozones.
Conodonts recovered from shell beds of the ‘Suri’ Formation in the Chaschuil
region indicate the B. navis Biozone (Albanesi and Vaccari, 1994), while
conodonts from the central Famatina span from the P. elegans to the P.
originalis Zones (Lehnert et al., 1997). The brachiopod-bearing
coquines from the upper Suri and lower Molles formations have yielded slightly
younger conodonts (Oepikodus evae Zone) (Albanesi and Astini, 2000b).
Chronology
of Tremadocian faunas of the Central Andean basin has not been accurately
established. In the last years, however, the classical trilobite biozonation
proposed by Harrington and Leanza (1957) has been substantially improved on the
basis of conodont data (Rao and Hünicken, 1995; Rao and Flores, 1997; Rao,
1999; Tortello and Rao, 2000 and references therein). On the other hand,
graptolites are by far the most important biostratigraphic group to calibrate
the latest Tremadocian and Arenig successions (Toro, 1994, 1997; Albanesi et
al., 2001, amongst others).
Acritarch
and palynological assemblages have also been used to refinate the
biostratigraphic schema (Ottone et al., 1992, 1999; Rubinstein and Toro,
1999)
Biostratigraphic
succession
Precordillera
basin
1)
The lowermost currently known brachiopod association comes from the coalesced
spongereceptaculitid reef-mounds developed near the base of the San Juan
Formation (Cañas and Carrera, 1993). Only fragmentary specimens of a small
unidentified pentamerid have so far been recovered (Benedetto et al., in
press). Intensive sampling from these levels has yet to be carried out. Acording
to the conodont assemblage recovered from the La Silla section, these levels may
be correlative to the P. deltifer Zone of Baltoscandia (Lehnert, 1993,
1995; Albanesi et al., 1995b).
2)
Archaeorthis sanroquensis Zone. (= Archaeorthis Zone defined by
Herrera and Benedetto, 1991).
This
is the lowermost brachiopod zone formally recognized in the Precordilleran
carbonate platform. It is characterized by low-diversity assemblages dominated
by Archaeorthis sanroquensis Benedetto, a species having great spacial
continuity. Intensive sampling over the past three years in the Cerro La Silla,
Cerro San Roque and Cumillango sections substantially increased our knowledge on
taxonomic composition and stratigraphic ranges of brachiopods from the A.
sanroquensis Zone (Cech, 2000; Esbry, 2001; Benedetto, 2001a, in press b;
Benedetto et al., in press). Based on faunal variations through the zone,
a number of ‘assemblages’ have been recognized in a previous work (Benedetto
et al., in press). It should be noted that such a compositional changes
appear to have been linked to the gradual deepening of the basin. Lithological
and paleoecological evidence indicates a transition from high-energy,
shallow-water settings punctuated by storm events at the base to a relatively
more stable, inner shelf environment upward in the biozone. According to Cañas
(1999) the maximum flooding was reached near the boundary between the Archaeorthis
sanroquensis and Huacoella radiata zones.
The
following successive sub-assemblages, whose ranges may be equivalent to
‘sub-zones’, have been recognized within the A. sanroquensis Zone, in
ascending order: (a) the lowermost subassemblage, encompassing the lower 12-15 m
of the zone, is characterized by the abundance of the pentamerids Syntrophia
sanjuanina Benedetto and Parallelostrophia lasillaensis Benedetto;
(b) a very low diversity sub-assemblage in which pentamerids are nearly absent
and Orthidium prominens Benedetto and A. sanroquensis dominate;
(c) a third sub-assemblage containing a relatively more diverse fauna is marked
by the first appearance of Nothorthis marginata Benedetto, and the
tritoechiids Tritoechia (Parvitritoechia) preandina Benedetto,
Tritoechia (Tritoechia) prima Benedetto and other species
of Tritoechia not yet determined. Nothorthis termalis (Herrera and
Benedetto), recorded in the Agua Hedionda (Huaco) section, probably belongs to
this assembalage, but its precise stratigraphic position within the A.
sanroquensis Zone is in need of verification. The best stratigraphic
sections to study the Archaeorthis sanroquensis Zone are Cerro Cumillango
and Cerro La Silla, while at Agua Hedionda (core of the Huaco Anticline) and
Niquivil only its upper half is preserved.
(3)
Ranorthis niquivilensis Zone. The base of this zone is drawn at the first
apparition of the eponymous species, which occurs in beds immediately overlying
the silicified interval. R. niquivilensis is associated with entirely
calcified brachiopods, and persists approximately 10-15 m above the last
apparition of A. sanroquensis. The lower part of the zone is
distinguished by the co-occurrence of A. sanroquensis and R.
niquivilensis. Upward in the zone is noticeable the first apparition of
leptellinids, represented by Leptella (Leptella) variabilis Benedetto
and Herrera and other still poorly known species.
Associated
forms are Hesperonomia sp., Hesperonomiella sp., Paralenorthis sp.,
Orthidium sp., Tritoechia sp., and undetermined pentamerids. The
informal name ‘Ranorthis-Leptella’ assemblage may be provisionally
used to account for the distinctive co-occurrence of these genera in the upper
part of the R. niquivilensis Zone. This assemblage fills the gap between
the last apparition of Archaeorthis and the first apparition of Huacoella
radiata.
(4)
Huacoella radiata Zone (= Huacoella Zone of Herrera and Benedetto,
1991). Huacoella
radiata Benedetto and Herrera is a widespread leptellinid which
has been chosen to name this biozone. This species typically occurs in a 25-30 m
thick succession of burrowed skeletal wackestones deposited in a low-energy,
open marine environment below the fair-weather wave-base, punctuated by
sporadical storm events. Key species in this still relatively poorly known
assemblage include Hesperonomia cf. planidorsalis Ulrich and
Cooper, Orthidium cf. gemmiculum (Billings), Paralenorthis n.
sp.(not yet described), Leptella (Leptella) alata Benedetto
and Herrera, and Tritoechia gigas Benedetto and Punctolira? sp.
The H. radiata Zone is well exposed in the Cuesta de Huaco and Niquivil
sections.
(5)
Niquivilia extensa Zone (= Niquivilia Zone of Herrera and
Benedetto, 1991). This
zone occurs in a 30-35 m thick stratigraphic interval overlying the H.
radiata Zone. In the Niquivil section -one of the best exposures of this
zone- a small onlap between the stratigraphic ranges of Huacoella radiata and
Niquivilia extensa has been detected. Typical associated brachiopods are Hesperonomiella
sulcata Benedetto and Leptella (Leptella) cf. alata.
Other species sporadically present are Trondorthis? sp., Hesperonomia?
n. sp., Orthidium sp., Notoscaphidia sp., Tritoechia sp., Leptella
(Leptella) plana Benedeto and Herrera , Leptella (Leptella)
costellata Benedetto and Herrera, and Leptella (Petroria)
sp.
(6) Monorthis cumillangoensis Zone (=Monorthis Zone of Herrera and Benedetto, 1991). Everywhere in the basin this zone developed in agitated shallow water settings linked to the second episode of reef-mound development. It occurs in a 50-60 m thick succession of bioclastic grainstones and stromatoporoid patch reefs (Cañas, 1999; Carrera et al., 1999, amongst others). A rather different, highly distinctive suite of brachiopods appears for the first time in this zone, including Ffynnonia spondyliformis Benedetto, Oligorthis zondaensis Benedetto, Polytoechia? sp. and Aporthophyla neumani Benedetto (Benedetto, 2001a). M. cumillangoensis, however, is absolutly dominant and often occurs
as
monospecific concentrations which may indicate sorting and accumulation in
relatively shallow envirnonment. Other associated taxa, such as Orthidium sp.,
Leptella (Leptella) sp. and Paralenorthis sp., come from
the underlying zones.
Biostratigraphic
evidence demonstrates that the transition from carbonate ramp facies to basinal
black shales is clearly diachronous (Hünicken, 1985; Albanesi et al.,
1998b). Such a diachronism, which becomes evident since the late Arenig, results
from the development of a homoclinal ramp inclined northward (Cañas, 1999). In
the depocenter located in the Guandacol area the trasitional beds consist of a
ritmic succession of bioclastic micrograded wackestones and black mudstones
belonging to the Tripodus laevis Zone (Albanesi et al., 1999),
which correlates with the Monorthis cumillangoensis Zone (and probably
with the upper part of the Niquivilia extensa Zone). Brachiopods from
these levels have not yet investigated in detail. Some preliminary
identifications include Tritoechia sp., Orthidium sp., Leptella
(Petroria) sp., Leptella (Leptella) sp., Taffia?
sp., and Productorthis? sp.
(7)
Ahtiella argentina Zone (= Ahtiella Zone of Herrera and Benedetto,
1991). This
zone is separated from the preceeding zone by a 50-60 m thick succession of
scarcely fossiliferous bioturbated wackestones containing a low-diversity
brachiopod fauna dominated by Paralenorthis vulgaris Herrera and
Benedetto and Petroria rugosa (the Petroria-Paralenorthis Biofacies
recognized by Carrera et al., 1999). The uppermost 30-35 m of the San
Juan Formation consists of dark-gray burrowed bioclastic wackestones, often with
nodular stratification, bearing a highly distinctive brachiopod assemblage
formed, besides the index fossil of the zone, by Sanjuanella plicata Benedetto
and Herrera, Inversella (Reinversella) arancibiai Herrera
and Benedetto, Taffia anomala Benedetto and Herrera, Productorthis
cienagaensis Herrera and Benedetto, Rugostrophia sp. (probably new,
not yet described), Camerella sp., Idiostrophia sp., and several
species of Ffynnonia, Tritoechia and Martellia. Brachiopods
from the Ahtiella argentina Zone have been described by Benedetto, 1987;
Benedetto and Herrera, 1986a, 1986b, 1987a, 1987b; Herrera and Benedetto, 1987,
1989. An oustanding feature of this zone is that species of Ffynnonia, Tritoechia,
Martellia and probably Sanjuanella and Productorthis, are
different in the northern and southern portions of the basin. As stated above,
no evident changes in facies associations are recorded in the Precordilleran
carbonate shelf until the late Arenig, when an homoclinal ramp with a depocenter
located to the north developed (Cañas, 1999). By the early Llanvirn, lithologic
and paleoecologic evidence from the southern Precordillera sections
(e.g.Villicum, Talacasto) indicates a shallow subtidal environment in a mid-ramp
setting sporadically affected by storm-wave base
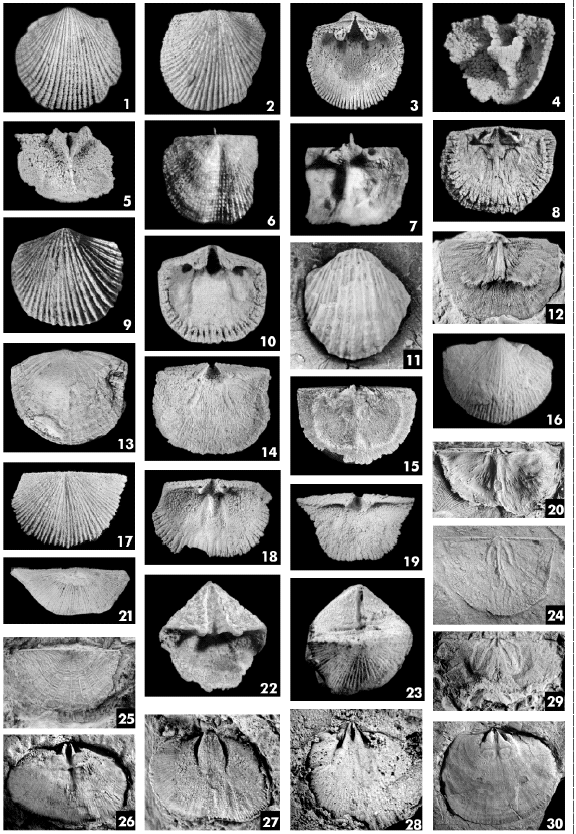
PLATE
1.
Figs.
1-3. Archaeorthis sanroquensis Benedetto. 1, exterior of ventral valve,
CEGH-UNC 17825, x2; 2, exterior of dorsal valve, CEGH-UNC 17824, x2; 3, interior
of ventral valve, CEGH-UNC 17801, x2.
Figs.
4, 5. Syntrophia sanjuanina Benedetto. 4, interior of ventral valve,
CEGH-UNC 20867, x 1,6; 5, interior of dorsal valve, CEGH-UNC 20866, x1,4.
Figs.
6, 7. Orthidium prominens Benedetto. Exterior and interior of dorsal
valve, CEGH-UNC 20776, x6,5.
Figs.
8-10. Nothorthis marginata Benedetto. 8, interior of dorsal valve,
CEGH-UNC 20745, x4; 10, exterior and interior of ventral valve, CEGH-UNC 30745,
x4.
Fig.
11. Ranorthis cumillangoensis Benedetto, exterior of ventral valve,
CEGH-UNC
Fig.
12. Huacoella radiata Benedetto and Herrera, interior of dorsal valve,
CEGH-UNC 175, x1,2.
Figs.
13-15. Niquivilia extensa Benedetto and Herrera. 13, exterior of ventral
valve, CEGH-UNC 210, x1,8; 14, interior of ventral valve, CEGH-UNC 172, x3; 15,
interior of dorsal valve, CEGH-UNC 227, x2.
Fig.
16. Leptella (Leptella) costellata Benedetto and Herrera, exterior of
ventral valve, CEGH-UNC 5115, x2.
Figs.
17-19. Monorthis cumillangoensis Benedetto. 17, exterior of dorsal valve,
CEGH-UNC 17924, x1,6; interior of dorsal valve, CEGH-UNC 17935, x 1,6; interior
of ventral valve, CEGH-UNC 17932, x1,5.
Figs.
20, 21. Ahtiella argentina Benedetto and Herrera, 10, interior of dorsal
valve, latex cast, CORD-PZ 8295, x3; exterior of ventral valve, CORD-PZ 8283,
x2.
Figs.
22, 23. Skenidioides n.sp., 22, interior of ventral valve, CEGH-UNC
20951, x4; conjoined specimen, dorsal view, CEGH-UNC 20952, x3,5.
Figs.
24, 25. Anchoramena cristata Benedetto. 24, internal view of dorsal valve,
CEGH-UNC 13686a, x1,5; 25, exterior of dorsal valve, CEGH-UNC 13695, x2.
Figs.
26, 27. Destombesium argentinum Benedetto. 26, internal mold of dorsal
valve, CEGH-UNC 17471, x1,5; 27, internal mold of ventral valve, CEGH-UNC 17465,
x2,3.
Fig.
28. Trucizetina chanarensis Benedetto. Internal mold of dorsal valve,
CEGH-UNC 17539, x2.
Fig.
29. Anisopleurella cf. gracilis (Jones). Internal mold of dorsal valve,
CORD-PZ 8238, x4.
Fig. 30. Hirnantia sagittifera (M’Coy). Internal mold of dorsal valve, CORD-PZ 8259, x 0,7
(Sánchez
et al., 1996; Carrera, 2001). The presence in these levels of abundant
demosponges, especially of large discoidal forms such as Patellispongia,
suggests soft or shifting substrates (Carrera, 1994, 1997). Conversely, in the
sections located to the north (Jáchal-Cerro Viejo area) the dominance of
elongated sponges and plectambonitoid brachiopods along with their occurrence in
nodular wakestones and marlstones are indicative of low-energy, deeper water
conditions. Hence, taxonomic differences between the northern and southern
portions of the basin can be attributed to an environmental gradient at the time
the Ahtiella argentina Zone developed (Sánchez et al., 1996).
(8)
Skenidioides assemblage. In the Cordón de las Chacritas the San Juan
Formation is conformably overlain by a 50 m thick succession of dark-gray
nodular limestones and marlstones assigned to the Las Chacritas Formation.
Compared to the preceeding Ahtiella argentina Zone, the Skenidioides assemblage
has in common several genera, such as Productorthis, Sanjuanella, Ffynnonia,
Petroria and Ahtiella, but all of them are represented by
different species (not yet described). One of the most distinctive taxa is Skenidioides
n.sp., which makes its first appearance in this assemblage. Moreover, this
species is always abundant at all localities in which this faunal association
has been reported.
Among
the plectambonitoids, Leptellina n. sp. and Benignites? n. sp.
(unpublished) are also restricted to this assemblage. The varied plectambonitoid
fauna, including the probably epiplantic Benignites (Havlicek, 1991),
suggests a deep-water, open-shelf environment. This is in agreement with the
dominance throughout the interval of the marginal-platform to upper-slope Periodon-Protopanderodus
conodont biofacies (Albanesi and Astini, 2000a). The Skenidioides assemblage
has also been recorded from many of the huge olistholiths embedded in the
Rinconada Formation exposed on the eastern slope of the Sierra Chica de Zonda.
The lower 50 m of the Las Aguaditas Formation are probably contemporary with the
Skenidioides assemblage, but this unit has not intensively been sampled
for brachiopods.
(9)
There are very few records of late Llanvirn (Llandeilo) brachiopods. Deposits of
that age have not yet been recorded in situ in the Precordillera basin,
with the exception of small outcrops in the
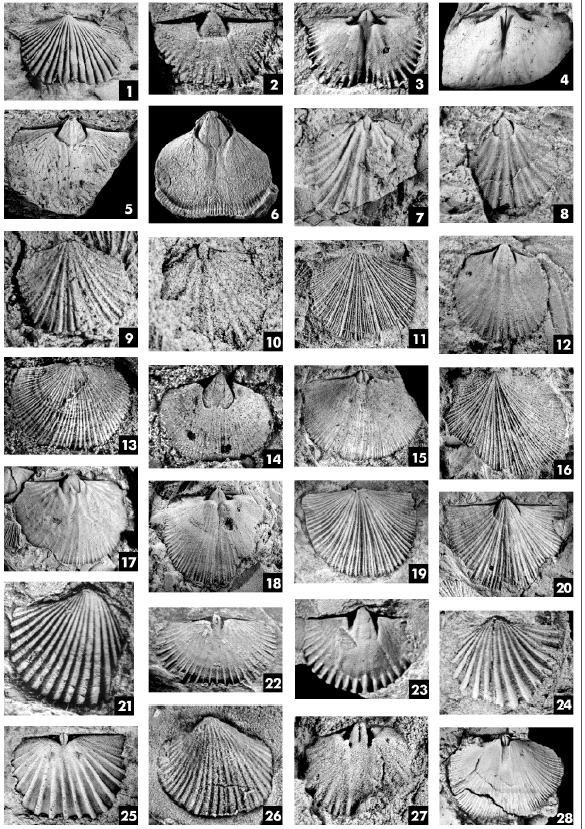
PLATE
2
Figs.
1-3. Suriorthis depressus Benedetto. Latex cast of ventral exterior,
CEGH-UNC 19029, x2,5; ventral internal mold of juvenile specimen CEGH-UNC 15919,
x5,5; internal mold of dorsal valve, CEGH-UNC 15762, x4.
Figs.
4, 6. Famatinorthis turneri Levy and Nullo. 4, internal mold of dorsal
valve CEGH-UNC 11004, x1; internal mold of ventral valve, CEGH-UNC 19700, x1,5.
Fig.
5. Mollesella planiventralis Benedetto. Internal mold of ventral valve,
CEGH-UNC 19649, x1.
Figs.
7, 8. Kvania n.sp.1. 7, internal mold of dorsal valve, CEGH-UNC 19465,
x7; 8, internal mold of ventral valve, CEGH-UNC 19466, x7. Figs. 9, 10. Kvania
n.sp.2. 9, latex replica of exterior, CEGH-UNC 19116, x4; 10, internal mold
of dorsal valve, CEGH-UNC 19470, x4.
Figs.
11, 12. Nanorthis n.sp. 11, latex cast of dorsal exterior, CEGH-UNC
18938, x3,5; 12, internal mold of ventral valve, CEGH-UNC 18934, x3.
Figs.
13-15. Lipanorthis santalaurae Benedetto. 13, látex cast of dorsal
exterior, CEGH-UNC 20463, x2; 14, internal mold of ventral valve, CEGH-UNC
204744, x2; 15, internal mold of dorsal valve, CEGH-UNC 20483, x3.
Figs.
16-18. Nanorthis purmamarcaensis Benedetto. 16, latex replica of dorsal
exterior, CEGH-UNC 18864, x2,5; 17, internal mold of ventral valve, CEGH-UNC
18810, x1,7; 18, internal mold of dosal valve, CEGH-UNC 18814, x2.
Figs.
19, 20. Nanorthis saltensis (Harrington). 19, latex replica of dorsal
exterior, CEGH-UNC 18820, x2; 20, internal mold of ventral valve, CEGH-UNC
18821, x2.
Figs.
21-23. Desmorthis segnis Havlicek and Branisa. 21, látex replica of
incomplete ventral exterior, CEGH-UNC 14411, x4,6; 22, internal mold of dorsal
valve, CEGH-UNC 14388, x4; 23, internal mold of ventral valve, CEGHUNC 14382,
x4.
Figs.
24, 25. Paralenorthis altiplanicus Benedetto. 24, látex replica of
ventral exterior, CEGH-UNC 13844b, x3,8; 25, internal mold of dorsal valve,
CEGH-UNC 13840, x3,5.
Figs.
26, 27. Dalmanella cf. salopiensis Williams. 26, látex replica of
ventral exterior, CEGH-UNC 21001, x3; internal mold of dorsal valve, CEGH-UNC
21002, x3.
Fig. 28. Drabovinella mojotoroensis Benedetto, Internal mold of dorsal valve, CEGH-UNC 17386, x1,5.
Ponon
Trehue area, near the San Rafael village, which are thought to represent the
southern edge of the Cuyania Terrane (Ramos, 1995). In the Sierra de Villicum,
the lowest 15 m of the La Cantera Formation consists of clast-supported and
matrix-supported conglomerates which include a relatively high percentage of
fossiliferous clasts containing reworked bryozoan zoecia and fragmented
brachiopod valves. The age of these clasts, according to their conodont content,
is early Llandeilo (Eoplacognathus lindstroemi, the uppermost subzone of
the P. serra Zone, Albanesi et al., 1995a). In general,
preservation of brachiopods is too poor for taxonomic purposes.
The
lower part of the Lindero Formation exposed in the Ponon Trehue area, south of
Mendoza province (Bordonaro et al., 1996), is almost coeval with the
fossiliferous clasts from the base of the La Cantera Formation (E.
reclinatus-E. lindstroemi subzones of the P. serra Zone, Lehnert et
al., 1998).
The
thin leveles of crinoidal limestone have yielded the brachiopods Ptychoglyptus
virginensis Cooper and Nugnecella rafaelensis Levy and Nullo (Levy
and Nullo, 1975). The latter is an enigmatic taxon which is in need of revision.
According to Williams and Harper (2000), the type specimens of N. rafaelensis
probably include different taxa.
(10)
The next interval corresponds to the latest Llanvirn-earliest Caradoc Pygodus
anserinus Zone.
It
is partially represented by the deep-water, carbonate slope-apron deposits of
the Las Aguaditas Formation (Keller et al., 1993; Astini, 1995). The
hemipelagic mustones and calcareous shales contain graptolites, conodonts,
trilobites and rare radiolarians. Strongly reworked shelly faunas are present
only in the gravity mass flow deposits, especially in the matrix of the lower
megabreccia horizon, which contains abundant specimens of Dinorthis sp.
and other forms not yet described.
(11)
Anchoramena assemblage. Beds of early Caradoc age containing a varied
brachiopod fauna are exposed in the northern Precordillera (Las Plantas
Formation; Benedetto, 1995) and in the Sierra de Villicum (La Pola Formation;
Benedetto in press c). Fossiliferous clasts embedded in the basal glacigenic
member of the Don Braulio Formation have also yielded approximately coeval
brachiopods (Benedetto, 1998b). The age of the Las Plantas Formation is well
constrained on the basis of graptolite content which is indicative of the C.
bicornis Zone (Ortega and Brussa, 1990; Albanesi and Ortega, 1998).
Graptolites recovered from the upper part of the La Pola Formation suggest a
similar age (Gisbornian to early Eastonian; Brussa, 2000). Despite taxonomic
composition of the three faunas is clearly different, they have in common at
least four genera, of which the Precordilleran endemic genus Anchoramena has
been chosen to nominate this assemblage. It should be noted that these faunas
include two environmentally different assemblages. The authochthonous fraction,
recorded in the deep-water black shales of the Las Plantas Formation, is
characterized by Oanduporella alamensis Benedetto, Bystromena? protegula
Benedetto and Sowerbyella cf. sericea (J. de C. Sowerby). The
genus Sowerbyella is also widespread in the hemipelagic facies of the
upper part of the Las Aguaditas Formation, in the upper member of the
Gualacamayo Formation (Benedetto et al., 1991) and in the slope facies of
the La Pola Formation, where is associated with Anoptambonites villicumensis Benedetto.
On
the other hand, brachiopods from the calcareous blocks of the Las Plantas
Formation, as well as those from the bioclastic sandstones of the La Pola
Formation (associated with bryozoans and solenoporacean algae), were
resedimented from shallower water settings located probably to the east of the
basin (Astini, 2001). These para-authochthonous or allochthonous concentrations
include thick-shelled rhynchonellids (Rostricellula robusta Benedetto),
orthids (Dinorthis polaensis Benedetto, Dinorthis cf. flabellum
(J. de C. Sowerby), Glyptorthis sp., Hesperorthis sp., Howellites
cf. macrostoma (Barrande), Tissintia robusta Benedetto, Campylorthis
gualcamayensis Benedetto, Onniella inversa Benedetto),
clitambonitoideans (Atelelasma sp.), triplesiids (three species of Bicuspina),
plecatambonitoideans (Aegiromena glacialis Benedetto), and
strophomenoideans (Oepikoides notus Benedetto, Anchoramena cristata Benedetto,
Glyptomena cf. sculpturata Cooper).
(12)
Destombesium assemblage. In the northern Precordillera the Anchoramena
assemblage is succeeded by a low-diversity brachiopod assemblage present in
the lower part of the overlying Trapiche Formation (Benedetto, 1999a). The fauna
was recovered from a 50-60 m thick interval of debris flows laying approximately
150 above the base of the unit. These beds contain Amorphognatus aff. superbus,
which suggests a late Caradoc-earliest Ashgill age (Albanesi et al.,
1995c). Three species are typical of this assemblage: Destombesium argentinum
Benedetto, Reuschella sp., and Rhynchotrema sp.
Geographycally, it is restricted to the Rio Blanco area, north of Jáchal
village. No other timeequivalent brachiopod faunas have been documented in the
Precordillera basin.
(13)
Trucizetina assemblage. The uppermost levels of the Trapiche Formation
contain a different assemblage dominated by Trucizetina chanarensis Benedetto
(Benedetto, 1999a). Preservation of associated brachiopods is not good enough to
make precise taxonomic identifications. Benedetto and Herrera (1987c) reported
from the same levels Holtedhalina? sp., Cyphomena? sp. and Dalmanella
aff. testudinaria, the latter re-assigned to T. chanarensis (Benedetto,
1999a). The age of this assemblage is not older than early Ashgill owing its
stratigraphic position well above to the Destombesium assemblage.
Although known species of Trucizetina from China and Bohemia are
Hirnantian in age, no typical elements of the Hirnantia Fauna have been
found together Trucizetina in the Trapiche Formation. It thus appears
that this assemblage may be late, but not latest, Ashgill in age.
(14)
Hirnantia sagittifera assemblage. It has long been known that shelly
faunas from the Don Braulio Formation exposed at the Sierra de Villicum are late
Ashgill in age (Levy and Nullo, 1974; Baldis and Blasco, 1975). This assemblage
includes most of typical elements of the widespread Hirnantia Fauna such
as Hirnantia sagittifera (M’Coy), Dalmanella testudinaria (Dalman),
Eostropheodonta hirnantensis (M’Coy), Plectothyrella crassicosta (Dalman),
Paromalomena polonica (Temple) and Cliftonia oxoplecioides Wright
(Benedetto, 1986, 1990). Leptaena trifidum Marek and Havlicek is also
present (unpublished). Drabovia undulata has only been reported from the
Cerro La Chilca outcrops (Astini and Benedetto, 1992).
(15)
Anisopleurella assemblage. A low diversity fauna composed by Anisopleurella
cf. gracilis (Jones) and Reuschella sp. has been recovered
from the fine-grained succession underlying the early Llandovery La Chilca
Formation at the Cerro del Fuerte (Benedetto et al., 1986b; Benedetto,
1987b). The latest Ashgill age of the Anisopleurella faunule is well
constrained on the basis of its association with Normalograptus persculptus.
However, its precise relationships with the above mentioned Hirnantia assemblage
are not clear. One interpretation of faunal differences between them is that the
Anisopleurella assemblage represents a deep-water lateral equivalent of
the Hirnantia Fauna (Sánchez et al., 1991).
Another
possibility is that the Anisopleurella fauna post-dates the Hirnantia assemblage.
This is supported by the fact that in the Don Braulio Formation N. perscultus
appears in the dark gray mudstones deposited just above the calcareous
siltstones bearing the Hirnantia Fauna, to which coexists in an interval
of 1-2 m. It thus appears that, as in southwest Wales (Cocks and Price, 1975), A.
gracilis is present in between the Hirnantia Fauna and the earliest
Silurian (Rhuddanian) fauna.
Sierra
de Famatina assemblages
The
early Ordovician volcano-sedimentary rocks of the Sierra de Famatina are
represented by the Suri and Molles formations, excelently exposed along the
Saladillo Grande, Cachiyuyo and Molles rivers in the central region of the
Famatina Range, La Rioja province. Less continuous but equally richly
fossiliferous successions crop out in the Chaschuil area, Catamarca Province
(Aceñolaza and Toselli, 1977; Mángano and Butois, 1996; Benedetto, 1994). Most
of the Famatinian brachiopods come from the bioturbated mudstones of the upper
member of the Suri Formation and the lower third of the overlying Molles
Formation, the latter formed by green pyroclastic breccias and volcanic
sandstones interbedded with tuffaceous fossiliferous siltstones (Astini and
Benedetto, 1996; Benedetto, in press a).
Three
successive brachiopod assemblages can be recognized in the Arenig
volcano-sedimentary rocks of the Sierra de Famatina.
(1)
Suriorthis depressus assemblage. The transition from the gray-bluish
laminated shales to the bioturbated greenish mustones of the upper part of the
Suri Formation is associated with the appearance of diverse brachiopod,
trilobite and bivalve faunas. Typical brachiopods from these levels are Paralenorthis
suriensis Benedetto, Crossiskenidium? stelzneri Benedetto, Protoskenidioides
cf. revelata Williams, Hesperonomia orientalis Benedetto, Suriorthis
depressus Benedetto, Hesperonomiella arcuata Benedetto, Panderina?
ambigua Benedetto, Ffynnonia famatinensis Benedetto, Rugostrophia
protoandina Benedetto, and Trigonostrophia reversa Benedetto
(Benedetto, in pess a).
(2)
Mollesella planiventralis assemblage. It occurs in the upper half of the
Molles Formation, approximately 500 m above the Suriorthis depressus assemblage.
The assemblage starts with lowdiversity coquines dominated by Paralenorthis
riojanus (Levy and Nullo), reaching its maximum diversity in the overlying
dark-gray bioturbated mudstones. Distinctive taxa are Tritoechia mollesensis Benedetto,
Skenidioides kayseri Benedetto, Monorthis transversa Benedetto, Productorthis
angulensis Benedetto, and Mollesella planiventralis Benedetto. The
co-occurrence of several of these taxa in the Loma del Kilómetro Member of the
Suri Formation (sensu Mángano and Buatois, 1992), in the Chachuil area,
suggests that the two faunas could be time-equivalent. In the latter locality,
however, Famatinorthis turneri is very abundant and Mollesella and
Productorthis are absent, probably due to environmental differences. Both
lithology and faunal content suggest that the Chaschuil fossiliferous beds are
correlatable to the Molles Formation rather than the Suri Formation.
(3)
Some isolated outcrops lithologically comparable to the Molles Formation yielded
somewhat different brachiopod faunas. One of them, located to the north of the
Loma del Kilómetro locality, contains Ahtiella sp., Paralenorthis
riojanus and a different species of Famatinorthis. A comparable
association composed by Ahtiella, Paralenorthis and Skenidioides
was recorded in the Quebrada Honda (north of Rio Cachiyuyo section). On the
basis of morphologic changes through the Famatinorthis lineage
(unpublished) is believed that this fauna lies above to the Mollesella
planiventralis assemblage.
However,
no conodont data are still available to corroborate the age of these beds.
Puna
assemblages It is widely accepted that the Ordovician Puna
basin of northern Argentina evolved in a back-arc setting above an east-dipping
subduction zone (Bahlburg, 1990). This north-trending, strongly subsiding basin
accommodates more than 3,000 m of marine volcano-sedimentary rocks ranging in
age from early Tremadocian to Arenig. In the northern Puna they are overlain by
a turbiditic complex c.3,500 m thick reaching the Midlle Ordovician
(Llandeilo-early Caradoc?). Shelly faunas and microfossils are almost completely
absent, so age determinations have been based largely on graptolite faunas
(Bahlburg et al., 1990). Brachiopod and trilobite faunas are restricted
to the easternmost and westernmost parts of the basin. In the eastern belt, the
early Tremadocian Taique Formation exposed in the Sierra de Cobres has yielded Apheorthis
samensis, while the volcano-sedimentary Las Vicuñas Formation (early
Tremadocian, Moya et al., 1993; Vaccari and Waisfeld, 2000) cropping out
near the Argentina-Chile limit contains a new species of the genus Kvania (Benedetto,
2002). At Vega Pinato, located a few kilometers southward, a richly
fossiliferous volcano-sedimentary succession of Arenig age is exposed
(Koukharsky et al., 1996). The diverse brachiopod fauna includes Finkelnburgia
sp., Oligorthis? sp., Hesperonomia sp., Tritoechia sp.,
Pinatotoechia acantha Benedetto, Punasptrophia multiseptata Benedetto,
Jaanussonites? sp., Imbricatia sp., and Rugostrophia sp.
(Benedetto, 2001b). The Vega Pinato assemblage shares four taxa (44%) with the Suriorthis
depressus assemblage of the Famatina basin, including the distinctive
spinose tritoechiid P. acantha, which until now is restricted to these
two regions. This faunal affinity strongly supports the hypothesis that the
western Puna volcanics represent the northern continuation of the Famatina
volcanic belt (Benedetto, 1998a).
Cordillera
Oriental assemblages
The
southern part of the large Central Andean basin is superbly exposed in the
Cordillera Oriental of northwestern Argentina, where the early Ordovician
succession attains more than 3,500 m in thicknes. The stratigraphic
relationships between the numerous informal and formally defined formations have
not yet been completely elucidated because of lacking of basin-scale
stratigraphic analyses and comprehensive sedimentologic and paleontologic
studies. Turner (1964) recognized two main units, the Santa Rosita Formation
(latest Cambrian-Tremadocian) and the Acoite Formation (early-mid Arenig), both
forming the Santa Victoria Group. Moya (1999) redefined some infomal units
erected by Harrington and Leanza (1957) and recognized along the western slope
of the Cordillera Oriental (from lowest to highest) the Lampazar, Cardonal,
Saladillo and Parcha formations.
In
the eastern part, the stratigraphic succession is formed by the San José,
Caldera, Floresta, Aspero and San Bernardo formations, which are approximately
coeval with the western units (Moya, 1999).
(1)
The lowest beds bearing brachiopods come from the Lampazar Formation, assigned
to the Parabolina (Neoparabolina) frequens argentina Zone. According to
conodont evidence, these beds are latest Cambrian to earliest Tremadocian in age
(Rao and Hünicken, 1995). The small giraldiellid Kavania n.sp.1
(Benedetto, 2002) forms locally monospecific assemblages, but in general
brachiopods are extremely rare in these beds.
(2)
The overlying sandstone beds (Cardonal Formation and equivalents units) yielded
several species of Nanorthis (not yet published) and a different finely
costellate species of Kvania (n.sp. 2). As so far as I aware, the
type material of Nanorthis putilliformis (Kobayashi) has been recovered
from these levels at the Parcha section. Both morphology and stratigraphic range
of this species are in need of revision. Nanorthis saltensis (Kayser),
another poorly known species, has also been recorded from beds probably
belonging to the Cardonal Formation. Preliminary collections from these sandy
beds show clear taxonomic differences among different localities, which could be
related to local environmental changes. However, until intensive sampling is
accomplished, a vertical succession of these assemblages cannot be entirely
rejected. These faunas span the uppermost part of the P. (N.) frequens
argentina Zone and the Kainella meridionalis Zone.
(3)
The uppermost part of the Cardonal Formation, well exposed in the Sierra de
Cajas (Quebrada Amarilla section), bears monotypic assemblages of a species of Nanorthis
characterized by its distinctive unequally multicostellate ornament (Nanorthis
n.sp. 2, Carrasco, 2001, unpubl.). These beds have been referred to the C.
angulatus Zone (Rao, 1999). A rather similar form occurs at the top of the
Angosto del Moreno section (Moya and Monteros, 2000).
(4)
The brachiopod composition of the lower part of the late Tremadocian succession
(Conophrys minutula/ Bienvillia tetragonalis Zone, Floresta
Formation) is still poorly known. These fine-grained beds contain one or more
species of Nanorthis, Euorthisina sp. nov., and Lipanorthis
santalaurae Benedetto (Carrasco, 2001, unpubl., Benedetto and Carrasco, in
press).
(5)
Nanorthis purmamarcensis assemblage. The late Tremadocian beds (Notopeltis
orthometopa Zone, Aspero and Coquena formations) contain a more diverse
brachiopod association characterized by Nanorthis purmamarcensis Benedetto,
Nanorthis brachymyaria Benedetto, Lipanorthis andinus Benedetto,
and Astraborthis quebradensis Benedetto (Benedetto and Carrasco, in
press). The species N. purmamarcenis, which has been chosen to nominate
this assemblage, is very abundant in the cross-bedded, stormdrived sandstones
cropping out in the Purmamarca, Santa Victoria and Cerro San Bernardo sections.
(6)
Nanorthis grandis assemblage. The succeeding early Arenig strata are
dominated by Nanorthis grandis (Harrington) which usually forms
monospecific assemblages. No associated brachiopos have been yet described. The
transitional beds in between the above mentioned storm-related sandstones and
the overlying fine-grained succession (referred to the Parcha, San Bernardo and
lower part of the Acoite formations) have yielded in some places graptolites of
the latest Tremadocian Hunnegraptus copiosus Zone (Albanesi et al. 2001).
These units, however, are mostly early Arenig in age.
(7)
Desmorthis segnis assemblage. Younger strata crop out along the
westernmost part of the Cordillera Oriental, where a continuous, more than 2,300
m thick coarsening-up succession is superbly exposed in the Los
Colorados-Espinazo del Diablo area. These successions have been matter of
several biostratigraphic, sedimentologic, paleoecologic and paleobiologic
studies (Waisfeld, 1993; Astini and Waisfeld, 1993; Sánchez and Waisfeld, 1995;
Toro, 1997; Benedetto, 1998c, amongst others). The lower half of this succession
is barren in shelly faunas. The graptolite association indicates the T.
akzharensis Zone. It is ovelain by about 700 m bearing graptolites of the B.
deflexus Zone (Toro, 1997) containig low-diversity brachiopod assemblages
dominated by Desmorthis segnis
Havlicek
and Branisa. This asemblage disappears before the first apparition of P.
altiplanicus. (8) Paralenorthis altiplanicus assemblage. A notably
more diverse fauna appears above the D. segnis assemblage. Besides the
eponymous species, it is composed by Glyptorthis cf. imbrex Havlicek
and Branisa, Hesperonomia orientalis Benedetto, Incorthis? sp., Pleurorthis?
sp., Euorthisina sp., Camerella sp., and Panderina? cf. ambigua
Benedetto. This assemblage falls within the midlle Arenig D. bifidus Zone
(Benedetto, 1998c).
(9)
The upper beds of the succession consist of pink to reddish cross-bedded
sandstones (the so-called Sepulturas Formation) interpreted by Astini (1994) as
deposited in an estuarine environment. The age of this unit is not well
constrained because of lack of conodonts and graptoplites. Brachiopods recovered
from these levels are Incorthis aff. marocana Mergl, Salopia?
lipanensis Benedetto. Some other forms have yet to be described.
(10)
Late Llanvirn (Llandeilo)-Caradoc brachiopod assemblages are relatively rare in
northwestern Argentina. At the top of the Los Colorados succession there is a
green-olive mudstone unit which overlies paraconformably early Ordovician
strata. The base of this fine-grained unit has been interpreted by Astini (1994)
as a regional flooding surface. According to Waisfeld (1996), trilobite evidence
supports the correlation of these levels with the early Caradoc Santa Gertrudis
Formation.
Brachiopods
have not been fully studied. Until now, Monorthis coloradoensis Benedetto,
Scaphorthis sp. and Dalmanella cf. salopiensis Williams
have been recognized in the Los Colorados area. In the Quebrada Chamarra, north
of the Los Colorados village, the resedimented boulders from the overlying
Silurian diamictite contain coquines of Dinorthis sp. (Benedetto and
Toro, 1996). Outside the Los Colorados area, Caradoc brachiopods are only known
from the Santa Gertrudis Formation, exposed in the Sierra de Mojotoro, which
contains a monotypic assemblage of Drabovinella mojotoroensis Benedetto,
1999a. Its early Caradoc age is supported by the conodont E. quadridactylus (Albanesi
and Rao, 1996). Outside Argentina, Drabovinella has been reported from
the Bolivian portion of the Central Andean basin, but since it is represented by
different species the correlation is uncertain (Havlicek and Branisa, 1980; Suárez
Soruco and Benedetto, 1996).
Aknowledgments.
Discussion about Ordovician biostratigraphy,
paleoenvironments and biofacies have been held with numerous workers of the
Paleobiology Group of the Cordoba University including B. Waisfeld, M. Carrera,
T.M. Sánchez, and N. Cech who provided invaluable discussion and comment. This
research was supported, in part, by a grant from the Agencia Nacional de
Promocion Cientifica y Tecnologica of Argentina (FONCYT, Grant PICT 5387).
References
Aceñolaza,
F.G. and Toselli, A.J., 1977. Observaciones geológicas y paleontológicas sobre el Ordovícico de
la zona de Chaschuil, Provincia de Catamarca. Acta Geologica Lilloana,
14:55-81.
Albanesi, G.L. and Astini, R.A., 2000a. Refined conodont biostratigraphy for Las Chacritas Formation, Argentine Precordillera. Ameghiniana 37 (4) supl.: 68R.
Albanesi,
G.L. and Astini, R.A., 2000b. New conodont fauna from Suri Formation
(Early-middle Ordovician), Famatina System, western Argentina. Ameghiniana
37 (4) supl.: Reunión
Anual Comunicaciones Asociación Paleontológica Argentina, Abstracts, 3.
Albanesi,
G.L. and Ortega, G., 1998. Conodont and graptolite faunas from the Las Plantas
Formation and equivalent units (Caradoc) in the Argentine Precordillera. 7th
International Conodont Symposium, Abstracts:
1-2.
Albanesi,
G.L. and Rao, I.R., 1996. Conodont fauna from the Santa Gertrudis Formation
(Middle-Late Ordovician), Eastern Cordillera, northwestern Argentina. 6th
European Conodont Symposium ECOS VI,
Abstracts, 3.
Albanesi,
G.L. and Vaccari, N.E., 1994. Conodontos del Arenig en la Formación Suri, Sistema de Famatina,
Argentina. Revista Española de Micropaleontología, 26(2):125-146.
Albanesi,
G.L. Benedetto, J. L., and Gagnier, P.G., 1995a. Sacabambaspis
janvieri (Vertebrata)
y conodontes del Llandeiliano temprano en la Formación La Cantera,
Precordillera de San Juan, Argentina. Boletín de la Academia Nacional de
Ciencias, 60: 519-544.
Albanesi,
G.L., ¨Hünicken, M., and Ortega, G., 1995b. Review of Ordovician
conodont-graptolite biostratigraphy of the Argentine Precordillera. In: Cooper,
J.D., Droser, M.K. and Finney, S.C. (eds.) Ordovician Odyssey, Society for
Sedimentary Geology (SEPM) 77: 31-36.
Albanesi,
G.L., Hünicken, M., and Ortega, G., 1995c. Amorphognathus aff. superbus (Conodonta) en la Formación
Trapiche (Ordovícico Superior), Cerro Potrerillo, Precordillera Argentina. Boletín
de la Academia Nacional de Ciencias, 60: 461-468.
Albanesi,
G.L., Hünicken, M.A., and Barnes, C.R., 1998a. Bioestratigrafía de conodontes
de las secuencias ordovícicas del Cerro Potrerillo, Precordillera central de
San Juan, R. Agrneitna. Actas de la Academia Nacional de Ciencias, 12:
7-72.
Albanesi,
G.L., Ortega, G., and Hünicken, M.A., 1998b. High resolution
conodont-graptolite biostratigraphy controlling the diachronous boundary between
the San juan and Gualcamayo formations (Arenig-Llanvirn) in the Argentine
Precordillera. 7th International
Conodont Symposium, Abstracts: 2-4.
Albanesi,
G.L., Ortega, G., Barnes, C.R., and Hünicken, M.A., 1999. Conodont-graptolite
biostratigraphy of the Gualcamayo Formation (Middle Ordovician) in the
Gualcamayo-Guandacol rivers area, Argentina Precordillera. In: Kraft, P. and
Fatka, O. (eds.), Quo vadis Ordovician? Acta Universitatis Carolinae,
Geologica,
43 (1-2): 45-48.
Albanesi,
G.L., Ortega, G., and Zeballo, G., 2001. Late Tremadocian conodont-graptolite
biostratigraphy from NW Argentine basins. The Guide Book, abstracts &
Ordovician Silurian correlation Chart for the joint field meeting IGCP 410 and
IGCP 421, Mongolia: 121-123.
Astini,
R.A., 1994. Interpretación estratigráfica de la Formación Sepulturas (Ordovícico
inferior) y unidades análogas del Noroeste argentino: La Aloformación
Sepulturas. 5º Reunión Argentina de Sedimentología, pp. 9-15.
Astini,
R.A., 1995. Sedimentología de la Formación Las Aguaditas (talud carbonático)
y evolución de la cuenca precordillerana durante el Ordovícico medio. Revista
de la Asociación Geológica Argentina, 50: 143-164.
Astini,
R.A., 1999. Sedimentary
record, volcano-tectonic cyclicity and progressive emergence of an Early
Ordovician perigondwanan volcanic arc: the Famatina System, p.115-118. In: P.
Kraft and Fatka, O. (eds.), Quo vadis Ordovician? Acta
Universitatis Carolinae, Geologica,
43 (1/2): 115-118.
Astini,
R.A., 2001. La Formación La Pola (Ordovícico Superior): relicto erosivo de la
glaciación hirnantiana en la Precordillera Argentina. Revista de la Asociación
Geológica Argentina, 56 (3):
Astini,
R.A. and Benedetto, J.L., 1992. El Ashgilliano tardío (Hirnantiano) del Cerro La Chilca,
Precordillera de San Juan. Ameghiniana,
29 (2): 249-264.
Astini,
R.A. and Benedetto, J.L. 1996. Paleoenvironmental features and basin evolution
of a complex volcanic arc region in the proto-Andean western Gondwana. 3rd.
International Symposium on Andean Geodynamics, 755-758.
Astini,
R.A. and Waisfeld, B.G., 1993. Análisis estratigráfico y paleoambiental del Ordovícico Inferior
(Formaciones Acoite y Sepulturas) al oeste de Purmamarca, Cordillera Oriental
argentina. 12º Congreso Geológico Argentino y 2º Congreso de Exploración
de Hidrocarburos, 1: 96-106
Astini,
R.A., Benedetto, J.L. and Vaccari, N.E., 1995. The Early Paleozoic evolution of
the Argentine Precordillera as a rifted, drifted and collided terrane: a
geodynamic model. Geological Society of America, Bulletin 107: 253-273.
Bahlburg,
H., 1990. The Ordovician basin in the Puna of NW Argentina and N Chile:
geodynamic evolution from back-arc to foreland basin. Geotektonische
Forschungen 75, p. 1-107
Bahlburg,
H., Maletz, J., Moya, M.C., and Salfity, J.A. 1990. The Ordovician sedimentary
rocks in the northern Puna of Argentina and Chile: new stratigraphical data
based on graptolites. Newsletter on Stratigraphy 23: 68-89.
Baldis,
B.A. and Blasco, G., 1975. Primeros trilobites Ashgillianos del Ordovícico sudamericano. 1er
Congreso Argentino de Paleontología y Bioestratigrafía, 1: 33-48.
Benedetto,
J.L, 1986. The first typical Hirnantia Fauna from South America (San Juan
Province, Argentine Precordillera). In: Racheboeuf, P.R. and Emig, D. (eds.),
Les Brachiopodes fossiles et actuels, Biostratigraphie du Paleozoïque,
4: 439-477.
Benedetto,
J.L., 1987a. Braquiópodos
clitambonitáceos de la Formación San Juan (Ordovícico temprano),
Precordillera de San Juan, Argentina. Ameghiniana,
24 (1-2): 95-108.
Benedetto,
J.L., 1987b. Braquiópodos
neo-ordovícicos del flanco occidental del Cerro del Fuerte, Provincia de San
Juan. Ameghiniana, 24 (3-4):169-174.
Benedetto,
J.L., 1990. Los géneros
Cliftonia y Paromalomena (Brachiopoda) en el Ashgilliano tardío
de la Sierra de Villicum, Precordillera de San Juan. Ameghiniana,
27 (1-2): 151-159.
Benedetto,
J.L., 1993. La hipótesis
de la aloctonía de la Precordillera Argentina: un test estratigráfico y
biogeográfico. 12º Congreso Geológico Argentino, Actas 3: 375-384.
Benedetto,
J.L., 1994. Braquiópodos
ordovícicos (Arenigiano) de la Formación Suri en la región de Chaschuil,
Sistema de Famatina, Argentina. Ameghiniana,
31 (3): 221-238.
Benedetto,
J.L., 1995. La fauna
de braquiópodos de la Formación Las Plantas (Ordovícico tardío, Caradoc),
Precordillera Argentina. Revista Española de Paleontología, 10 (2):
239-258.
Benedetto,
J.L., 1998a. Early Palaeozoic brachiopods and associated shelly faunas from
western Gondwana: its bearing on the gedynamic history of the pre- Andean
margin. In: Pankhurst,R. & Rapela, C.W. (eds.), The proto-Adean margin of
Gondwana, Geological Society London, Special Publications, 142: 57-83.
Benedetto,
J.L., 1998b. Braquiópodos
Caradocianos en los bloques de la diamictita glacigénica de la Formación Don
Braulio (Ashgilliano), Sierra de Villicum, Precordillera Argentina. Ameghiniana
35 (3): 243-254.
Benedetto,
J.L., 1998c. Early Ordovician (Arenig) brachiopods from the Acoite and
Sepulturas formations, Cordillera Oriental, northwestern Argentina. Geologica
et Palaeontologica,
32: 7-27.
Benedetto,
J.L., 1999a. Braquiópodos de la Formacion Trapiche, Ordovícico tardío de la
Precordillera de San Juan y La Rioja, Argentina. Ameghiniana,
36 (3): 327-341.
Benedetto,
J.L., 1999b. El genero
Drabovinella (Brachiopoda) en el Caradociano de las Sierra de Mojotoro,
Provincia de Salta, Argentina. Ameghiniana,
36 (2): 235-238.
Benedetto,
J.L., 2001a. Silicified early Ordovician (Arenig) brachiopods from the San Juan
Limestone, Argentine Precordillera. Geologica et Palaeontologica,
35: 1-29.
Benedetto,
J.L., 2001b. Una fauna
de braquiópodos arenigianos (Ordovícico temprano) en rocas volcaniclásticas
de la Puna occidental: Implicaciones paleoclimáticas y paleogeográficas. Ameghiniana,
38 (2): 131-146.
Benedetto,
J.L., 2001c. Palaeolatitudinal distribution patterns of higher rhynchonelliform
brachiopods in the early Ordovician. In: Brunton, C.H.C., Cocks, L.R.M. and
Long, S.L. (eds.), Brachiopods, past and present. The Systematics Association
Special Volume Series 63: 299-314, Taylor & Francis, London.
Benedetto,
J.L., 2002. El género
Kvania Havlicek en el Tremadociano Temprano del noroeste de Argentina y
su significado en la radiación basal de los braquiópodos plectorthoideos. 8º
Congreso Argentino de Paleontología y Bioestratigrafía. Resúmenes.
Benedetto,
J.L., in press a. Early Ordovician (Arenig) brachiopods from volcaniclastic
rocks of the Famatina Range, nothwest Argentina. Journal of Paleontology.
Benedetto,
J.L., in press b. El género Ranorthis Öpik (Brachiopoda) en el Ordovícico
Temprano de la Precordillera Argentina: Interés bioestratigráfico, filogenético
y paleobiogeográfico. Ameghiniana.
Benedetto,
J.L., in press c. Braquiópodos del Caradociano Temprano de la Formación La
Pola, Sierra de Villicum, Precordillera de San Juan (Argentina). Ameghiniana.
Benedetto,
J.L. in press d. Brachiopods. In: Benedetto, J.L. (ed.), Ordovician fossils of
Argentina. Chapter 7. Secretaría
de Ciencia y Tecnología, Universidad Nacional de Córdoba.
Benedetto,
J.L. and Astini, R.A., 1993. A collisional model for the stratigraphic evolution
of the Argentine Precordillera during the Early Paleozoic. 2nd International
Symposium on Andean Geodynamics, pp.501-504, Oxford. ORSTOM Ed.
Benedetto,
J.L. and Carrasco, P.A., in press. Tremadoc (earliest Ordovician) brachiopods
from the Purmamarca region and Sierra de Mojotoro, Cordillera Oriental of
northwestern Argentina. Geobios.
Benedetto,
J.L. and Herrera, Z.A., 1986. Braquiópodos del Suborden Strophomenidina de la Formación San Juan
(Ordovícico temprano), Precordillera de San Juan, Argentina. 4º Congreso
Argentino Paleontología y Bioestratigrafía, Actas 1: 113-123.
Benedetto,
J.L. and Herrera, Z.A., 1987a. Sanjuanella, un nuevo género de la Subfamilia Ahtiellinae
(Brachiopoda, Plectambonitacea) del Ordovícico de la Precordillera Argentina. 4º
Congreso Latinoamericano de Paleontología, Actas 1: 97-109.
Benedetto,
J.L. and Herrera,Z.A., 1987b. El género Platystrophia King (Brachiopoda) en la Formación
San Juan de la Precordillera Argentina. Ameghiniana,
24 (1-2): 51-59.
Benedetto,
J.L. and Herrera, Z.A., 1987c. Primer hallazgo de braquiópodos y trilobites en la Formación
Trapiche (Ordovícico tardío), Precordillera Argentina. 10º Congreso Geológico
Argentino, Actas 3: 73-76.
Benedetto,
J.L. and Herrera, Z.A., 1993. New Early Ordovician Leptellinidae (Brachiopoda)
from the San Juan Formation, Argentina. Ameghiniana, 30 (1):
39-58.
Benedetto,
J.L. and Sánchez, T.M., 1990. Fauna y edad del estratotipo de la Formación Salar del Rincón
(Eopaleozoico, Puna Argentina). Ameghiniana, 27 (3-4): 317-326.
Benedetto,
J.L. and Sánchez, T.M. , 1996. Paleobiogeography of brachiopod and molluscan
faunas along the South American margin during the Ordovician. In:
Baldis, B.A.y Aceñolaza,F.G., El Paleozoico inferior del Noroeste de Gondwana, Correlación
Geológica, 12: 23-38.
Benedetto,J.L.and
Toro, B.A., 1996. Hallazgo
de braquiópodos del Ordovícico tardío y Silúrico en la Cordillera Oriental
de Jujuy, Argentina. Ameghiniana,
33 (2): 228-229.
Benedetto,
J.L., Cañas, F.L., and Astini, R.A., 1986a. Braquiópodos y trilobites de la zona de transición entre
las formaciones San Juan y Gualcamayo, Provincia de La Rioja, Argentina. 4º
Congreso Argentino Paleontología y Bioestratigrafia, Actas 1: 103-111.
Benedetto,
J.L., Ortega, G., Brussa, E.D and Toro, B.A., 1986b. Estratigrafía y fauna de
la secuencia neo-ordovícica y llandoveriana del Río Escondido (flanco
occidental del Cerro del Fuerte), Provincia de San Juan. Primeras Jornadas
Geología de la Precordillera, p. 128-133.
Benedetto,
J.L., Herrera, Z.A.,and Ortega, G., 1991. Bioestratigrafía del Ordovícico en el flanco occidental
del Cerro Potrerillo, Provincia de San Juan. Revista Asociación Geológica
Argentina, 46 (1-2): 60-68.
Benedetto,
J.L., Vaccari, N.E., Carrera, M.G. and Sánchez, T.M., 1995. The
evolution of faunal provincialism in the Argentine Precordillera during the
Ordovician: New evidence and paleogeographic implications. In: Cooper, J.D.,
Droser, M.K. and Finney, S.C. (eds.) Ordovician Odyssey. The Pacific section
Society for Sedimentary Geology (SEPM) 77: 181-184.
Benedetto,
J.L., Sánchez, T.M., Carrera, M.G., Brussa, E.D., and Salas, M.J., 1999.
Paleontological constraints on successive paleogeographic positions of
Precordillera terrane during the Early Paleozoic. Geological Society of
America, Special Paper 336: 21-42.
Benedetto,
J. L., Brussa, E.D. and Pompei, J. 2002. El Ordovícico de la región de Susques-Huancar (Puna
oriental de Jujuy): Precisiones sobre su edad y significado estratigráfico. 20º
Congreso Geológico Argentino.
Benedetto,
J.L., Cech, N., and Esbry, C., in press. New Late Temadoc-Early Arenig
(Early Ordovician) silicified brachiopods from the lower part of the San Juan
Formatioin, Argentine Precordillera. Ameghiniana.
Bordonaro,
O., Keller, M., and Lehnert,O., 1996. Wel Ordovícico de Ponon Trehue en la Provincia de Mendoza
(Argentina): redefiniciones estratigráficas. 13º Congreso Geológico
Argentino y 2º Congreso Exploración de Hidrocarburos, 1: 541-550.
Brussa,
E.D. 2000. Una nueva
asociación de graptolitos caradocianos en la Formación La Cantera, sierra de
Villicum, Precordillera argentina. Ameghiniana 37: 7R.
Cañas,
F.L., 1999. Facies
and sequences of the Late Cambrian-Early Ordovician carbonates of the Argentine
Precordillera: A stratigraphic comparison with Laurentian platforms. In:
Ramos,V. and Keppie, D., Gondwana- Laurentia connections before Pangea. Geological
Society of America, Special Paper 336: 43-62.
Carrasco,
P.A., 2001. Braquiópodos Ordovícicos (Tremadociano-Arenigiano temprano) del
noroeste de Argentina, con especial referencia a las especies del género Nanorthis.
Trabajo Final de Grado (Tesina), Escuela de Biología, Facultad de Ciencias
Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, 50 p. Unpubl.
Carrera,
M.G., 1994. An
Ordovician sponge fauna from the San Juan Formation, Precordillera basin,
western Argentina. Neus
Jahrbuch für Geologie und Pälaeontologie, Abh. 191: 201-221.
Carrera,
M.G., 1997.Significado paleoambiental de los poríferos y briozoos de la Formación
San Juan (Ordovícico), Precordillera argentina. Ameghiniana 34: 179-199.
Carrera,
M.G., 2001. Análisis de la distribución y composición de las biofacies de la
Formación San Juan (Ordovícico Temprano), Precordillera argentina. Ameghiniana,
38 (2):169-184.
Carrera,
M.G., Sánchez, T.M., and Benedetto, J.L., 1999. Paleoenvironmental controls on
biofacies in the early Ordovician limestones of the Argentine Precordillera. In:
Kraft, P. and Fatka, O. (eds.), Quo vadis Ordovician? Acta
Universitatis Carolinae,
Geologica,
43 (1-2): 475-478.
Cech,
N., 2000. Análisis paleoecológico y dinámica de las comunidades en el
Ordovícico temprano de la Precordillera Argentina. Tesis de Grado
(Tesina), Escuela de Biología, Facultad de Ciencias Exactas, Físicas y
Naturales, Universidad Nacional de Córdoba, 66 p. Inédito.
Cocks,
L.R.M. and Price, 1975. The biostratigraphy of the Upper Ordovician and Lower
Silurian of South-West Dyfed, with comments on the Hirnantia Fauna. Palaeontology,
18: 703-724.
Esbry,
C., 2001. El género Arechaeorthis en la Formación San Juan, Ordovícico
inferior de la Precordillera Argentina. Trabajo Final de Grado (Tesina),
Escuela de Biología, Facultad de Ciencias Exactas, Físicas y Naturales,
Universidad Nacional de Córdoba, 66 p. Unpubl.
Harrington,
H.J., 1938. Sobre las
faunas del Ordoviciano inferior del Norte Argentino. Revista del Museo de La
Plata (nueva serie), 1, Sección Paleontología, pp. 109-289.
Harrington,
H.J. and Leanza, A.F., 1957. Ordovician trilobites of Argentina. University
of Kansas Special Publication 1:1- 276.
Herrera,
Z. A. and Benedetto, J.L., 1987. El género Reinversella (Brachiopoda) en el Ordovícico
temprano de la Precordillera Argentina. X Congreso Geológico Argentino,
Actas 3: 77-80.
Herrera,
Z.A. and Benedetto, J.L., 1989. Braquiópodos del Suborden Orthidina de la Formación San Juan (Ordovícico
temprano) en el área de Huaco-Cerro Viejo, Precordillera Argentina. Ameghiniana,
26
(1-2): 3-22.
Herrera,
Z.A. and Benedetto, J.L., 1991. Early Ordovician brachiopod faunas from the
Precordillera basin, Western Argentina: biostratigraphy and paleobiogeographical
affinities. In: MacKinnon, D.I. et al., (eds.) Brachiopods through time,
p. 283-301. Balkema Ed.
Havlicek,
V., 1991. Genus concept in the Aegiromeninae and Leptellininae
(Plectambonitoidea, Brachiopoda). Vestník Ustøedního ústavu geologického,
66: 151-162.
Havlicek,
V. and Branisa, L., 1980. Ordovician brachiopods of Bolivia. Rozpravy
Ceskoslovenské Akademie Ved, 90:1- 53.
Hünicken,
M. , 1995. Lower Ordovician conodont biostratigraphy in Argentina. Boletín
de la Academia Nacional de Ciencias,
56: 309-322.
Keller,
M., Eberlein, S., and Lehnert, O., 1993. Sedimentology of Middle Ordovician
carbonates in the Argentine Precordillera: evidence of regional relative
sea-level changes. Geologische Rundschau, 82: 362-377.
Koukharsky,
M., Torres Claro, R., Etcheverría, M., Vaccari, N.E. and Waisfeld, B.G. 1996. Episodios
volcánicos del Tremadociano y Arenigiano en Vega Pinato, Puna salteña,
Argentina. 13º Congreso Geológico Argentino, 5: 535-542.
Lehnert,
O., 1993. Bioestratigrafía de los conodontes Arenigianos de la Formación San
Juan en la localidad de Niquivil (Precordillera sanjuanina, Argentina) y su
correlación intercontinental. Revista Española de Paleontología, 8:
153-164.
Lehnert,
O., 1995. The Tremadoc-Arenig transition in the Argentine Precordillera. In:
Cooper, J.D., Droser, M.K. and Finney, S.C. (eds.), Ordovician Odyssey. The
Pacific section Society for Sedimentary Geology (SEPM) 77: 145-148.
Lehnert,
O. T., S. M. Bergström, and N. E. Vaccari. 1997. Arenig conodonts from the
Famatina Range, northwestern Argentina: faunal affinities and paleogeographic
implications. Abstracts 18th IAS
Regional European Meeting of Sedimentology, Gaea Heidelbergensis,
3:216-217.
Lehnert,
O., Bergtröm, S.M., Keller, M., and Bordonaro, O., 1998. Ordovician
(Darriwilian-Caradocian) conodonts from the San Rafael Region, west-central
Argentina: biostratigraphic, paleoecologic, and paleogeographic implications. Bollettino
della Societá Paleontologica Italiana, 37 (2-3): 199-214.
Levy,
R., and Nullo, F. 1973. Braquiópodos
Ordovícicos de la Sierra de Famatina (Formación Molles), Provincia de La
Rioja. Ameghiniana,
10:139151. Levy, R. and Nullo, F., 1974. La fauna del Ordovícico (Ashgilliano) de Villicun, San
Juan, Argentina (Brachiopoda). Ameghiniana,
11 (2):173-194.
Levy,
R. and Nullo, F., 1974. La
fauna del Ordovícico (Ashgilliano) de Villicun, San Juan, Argentina
(Brachiopoda). Ameghiniana,
11 (2):173-194.
Levy,
R. and Nullo, F., 1975. Braquiópodos
Ordovícicos de Ponon Trehue, Bloque de San Rafael (Provincia de Mendoza,
Argentina). 1er Congreso Argentino de Paleontología y Bioestratigrafía, 1:
23-32.
Levy,
R. and Nullo, F., 1980. Braquiópodos
de la Sierra de Famatina (Formación Suri), Provincia de La Rioja. 2º
Congreso Argentino de Paleontología y Bioestratigrafía, 1: 31-47.
Mangano,
M. G., and Buatois, L.A., 1992. Análisis genético de concentraciones fósilesen una secuencia
volcaniclástica de plataforma. Ameghiniana, 29 (2): 135-151.
Mangano,
M. G., and Buatois, L.A., 1996. Shallow marine event sedimentation in a volcanic
arc-related setting: The Ordovician Suri Formation, Famatina Range, northwest
Argentina. Sedimentary
Geology,
105:63-90.
Moya,
C., 1999. Ordovícico en los Andes del Norte Argentino. In: González Bonorino,
G., Omarini, E.R. and Viramonte, J. (eds.), Geología del Noroeste Argentino. Relatorio
14º Congreso geológico Argentino, 1: 134-152.
Moya,
M.C. and Monteros, J.A., 2000. El Angosto del Moreno (Cordillera oriental Argentina), un área clave
para analizar el límite Cámbrico-Ordovícico y la discordancia Iruya. 14º
Congreso Geológico Boliviano, pp. 142-147.
Moya,
M.C., Malanca, S., Hongn, F.D., and Bahlburg, H. 1993. El
Tremadoc temprano en la Puna Occidental argentina. 12º Congreso Geológico
Argentino, 2: 20-30. Mendoza.
Ortega,
G. and Albanesi, G.L., 1999. Graptolite biostratigraphy of the of the Gualcamayo
Formation (Middle Ordovician) at the Los Sapitos creek section, Argentina
Precordillera. In: Kraft, P. and Fatka, O. (eds.), Quo vadis Ordovician? Acta
Universitatis Carolinae,
Geologica,
43 (1-2): 49-52.
Ortega,G.
and Brussa, E.D., 1990. La
subzona de Climacograptus bicornis (Caradociano temprano) en la Formación
Las Plantas en su localidad tipo. Ameghiniana,
27: 281-288.
Ottone,
E., Toro, B. and Waisfeld, B.G., 1992. Lower Orodovician palynomorphs from the
Acoite Formation, Northwestern Argentina. Palynology, 16: 93-116.
Ottone,
E.G., Albanesi, G.L., Ortega, G., and Holfeltz., 1999. Palynomorphs, conodonts,
and associated graptolites from the Ordovician Los Azules Formation, Central
Precordillera, Argentina. Micropaleontology, 45 (3): 225-250.
Rao,
R.I., 1999. Los conodontes Cambro-Ordovícicos de la Sierra de Cajas y el
Espinazo del Diablo, Cordillera Oriental, República Argentina. Revista Española
de Micropaleontología, 31: 23-51.
Rao,
R. I. and Flores, F.J., 1997. Conodontes Ordovícicos (Tremadoc Superior) de la Sierra de Aguilar,
provincia de Jujuy, Rep. Argentina. Bioestratigrafía y tafonomía. Revista
Española de Micropaleontología, 31: 23-51.
Rao,
R.I. and Hünicken, M., 1995. Conodont biostratigraphy of the
Cambrian-Ordovician boundary in northwestern Argentina. In: Cooper, J.D.,
Droser, M.K. and Finney, S.C. (eds.), Ordovician Odyssey. The Pacific section
Society for Sedimentary Geology (SEPM) 77: 125-128
Ramos,
V. ,1995. Sudamérica, un mosaico de continentes y océanos. Ciencia Hoy,
6(32): 24-29.
Rubinstein,
C.V. and Toro, B.A., 2001. Review of acritarch biostratigraphy in the Arenig of
the Eastern Cordillera, Northwestern Argentina: New data and calibrations with
graptoilite zonation. Contributions to Geology and Palaeontology of Gondwana
in honour of Helmut Wopfner Sánchez, T.M. and Waisfeld, B.G., 1995. Benthic
assemblages in the northwestern of Gondwana: A test of the Ordovician
evolutionary radiaton model In: Cooper, J.D., Droser, M.K. and Finney, S.C.
(eds.), Ordovician Odyssey. The Pacific section Society for Sedimentary
Geology (SEPM) 77: 409-412.
Sánchez,
T.M., Benedetto, J.L. and Brussa, E.D., 1991. Late Ordovician stratigraphy,
paleoecology and sea level changes in the Argentine Precordillera. In: Barnes,
Ch. and Williams, S.H. (eds.), Advances in Ordovician Geology, Geological
Survey of Canada, Paper 90-9: 245-258.
Sánchez,
T.M., Carrera, M. and Benedetto, J.L., 1996 Variaciones faunísticas en el techo
de la Formación San Juan (Ordovícico temprano, Precordillera Argentina):
significado paleoambiental. Ameghiniana,
33
(2): 185-200.
Suárez
Soruco, R. and Benedetto, J.L., 1996. Primeros datos sobre la fauna de braquiópodos de la
Formación San Benito en la Cordillera del Tunari, Cochabamaba, Bolivia. Actas
12º Congreso Geológico Boliviano, I: 211-216.
Toro,
B. A., 1994. Las zonas de Didymograptus (Didymograptellus) bifidus (Arenigiano
medio) y Didymograptus (Corymbograptus) deflexus (Arenigiano
inferior) en la Formación Acoite, Cordillera Oriental, Argentina. Ameghiniana,
31: 209-220.
TORO, B. A., 1997. La fauna de graptolitos del la Formación Acoite en el
borde occidental de la Cordillera Oriental Argentina. Parte I: Análisis
bioestratigráfico. Ameghiniana,
34: 393-412.
Toro,
B. A. and Brussa, E.D., 1997. Graptolitos de la Formación Suri (Arenig) en el Sistema de Famatina,
Argentina. Revista Española de Paleontología, 12(2): 175-1884.
Tortello,
M .F. and Rao, R.I., 2000. Trilobites y conodontes del Ordovícico temprano del Angosto de
Lampazar (provincia de Salta, Argentina). Boletín Geológico y Minero, 111
(2-3): 61-84.
Turner,
J.C.M., 1964. Descripción
geológica de la Hoja 2c, Santa Victoria, (provincias de Salta y Jujuy). Boletín
del Instituto Nacional de Geología y Minería, 104: 1-83.
Vaccari,
N. E. and Waisfeld, B.G., 1994. Nuevos trilobites de la Formación Suri (Ordovícico temprano) en la
región de Chaschuil, Provincia de Catamarca: Implicancias bioestratigráficas. Ameghiniana,
31(1):73-86.
Vaccari,
N.E. and Waisfeld, B.G., 2000. Trilobites Tremadocianos de la Formación Las Vicuñas, Puna
occidental, Provincia de Salta, Argentina. Memorias 14º Congreso Geológico
Boliviano, p. 165-169.
Vaccari,
N.E., Benedetto, J.L., Waisfeld, B.G. and Sánchez, T.M., 1993. La
fauna de Neseuretus en la Formación Suri (Oeste de Argentina): edad y
relaciones paleobiogeográficas. Revista Española de Paleontología, 8
(2): 185-190.
Waisfeld,
B.G., 1993. Concentraciones
fosilíferas autóctonas y para-autóctonas en la Formación Acoite (Ordovícico
temprano), Cordillera Oriental, Provincia de Jujuiy. Consideraciones tafonómicas
y paleoecológicas. 12º Congreso Geológico Argentino, 2: 296-303.
Waisfeld,
B.G., 1996. Revisión
de la Zona de ‘Hoekaspis schalagintweiti’ Harrington & Leanza,
Ordovícico del Noroeste de Argentina. 12º Congreso Geológico de Bolivia,
2: 915-922.
Williams,
A. and Harper, D.A.T., 2000. Orthida. In: Kaesler, R.L. (ed.), Treatise on
Invertebrate Paleontology, Part H, Brachiopoda (revised), 3: 714-844. The
Geological Society of America and University of Kansas Press,
Recibido:
18 de Septiembre
de 2002