Ordovician
Palynomorphs of Argentina: An Integrated Approach
Maria
del Milagro VERGEL1, Lucia ARAOZ2 and
Claudia V. RUBINSTEIN3
1 CONICET / Facultad de Ciencias Naturales e I.M.L. (UNT). Miguel Lillo 205 – San Miguel de Tucumán, Argentina. E-mail: vergelmar@tucbbs.com.ar
2 INSUGEO – CONICET. Miguel Lillo 205, 4000 San Miguel de Tucuman. Argentina. E-mail: insugeo@unt.edu.ar
3 CONICET - IANIGLIA – CRICYT. cc: 131- 5500 Mendoza, Argentina. E-mail: crubinstein@arlinkbbt.com.ar
Abstract:
ORDOVICIAN PALYNOMORPHS OF ARGENTINA:
AN INTEGRATED APPROACH.
The
knowledge of Ordovician Argentine palynomorphs increased substantially in the
last decade. The temporal distribution of them (acritarchs, chitinozoans,
scolecodonts, prasinophycean algae and cryptospores) embraces the entire
Ordovician, from the early Tremadocian to the latest Ordovician (post
Hirnantian) - Asghill. At the moment, they have only been documented in the
Cordillera Oriental, Precordillera, Famatina and Puna Basins. Biostratigraphic
and paleogeographic data with paleoecological inferences are summarized.
Resumen:
PALINOMORFOS
ORDOVICICOS
DE ARGENTINA:
UNA VISION INTEGRADORA.
El conocimiento de
palinomorfos (acritarchs, chitinozoans, scolecodonts, prasinophycean algae and
cryptospores) ordovícicos de Argentina se ha incrementado en la última década.
Su distribución temporal abarca desde el Ordovícico temprano (Tremadociano
Inferior) al Asghiliano tardío. Hasta el momento, ellos han sido registrados en
Cordillera Oriental, Precordillera, Famatina y Puna. Se resumen datos
bioestratigráficos, paleogeográficos e inferencias paleoecológicas.
Key
words: Ordovician. Palinomorphs. Argentina.
Palabras
clave: Palinomorfos. Ordovicico.
Argentina.
Introduction
Although
palynological researches were unknown in the Ordovician rocks of Argentina
before 1970s, the situation dramatically changed during the following two
decades. The incentive was the recognized biostratigraphic potential of such
organic-walled microfossil group, mainly used to establish biostratigraphical
correlations, as well as the palaeobiogeographical and palaeoenvironmental
interpretation with applications in the field of geochemistry. The integrated
palynological studies with other important Ordovician fossil groups (trilobites,
graptolites and conodonts) highlighted its important applications.
In
the present report we considered different palynomorphs: acritarchs,
chitinozoans, scolecodonts, prasinophycean algae and cryptospores, all of them
with record in the Ordovician strata of Argentina.
Due
to the scarcity of contributions related to Ordovician palynomorpohs from
Argentina, this synthesis is organized by geographic regions and stratigraphic
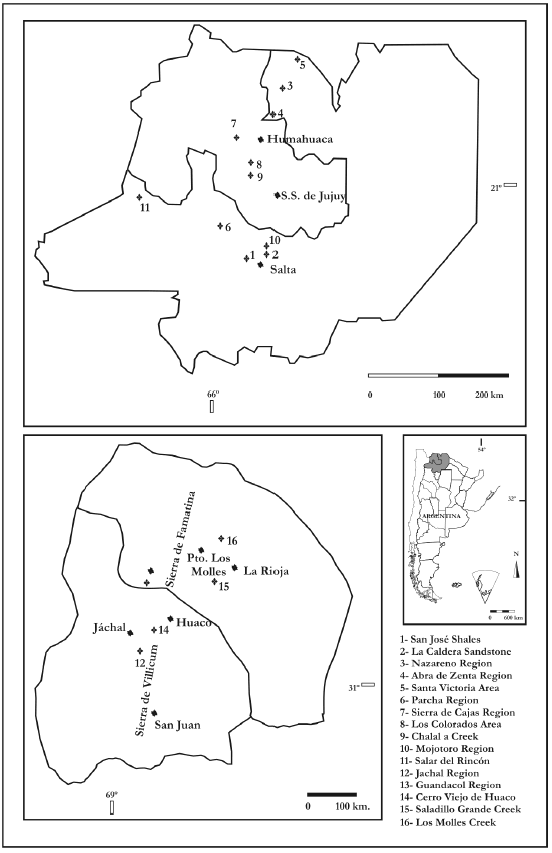
ranges. The geographic location of records are shown in figure 1. Plates that
illustrates this article have been extracted from the original papers cited in
the text.
At
the moment Ordovician palynomorphs from Argentina have only been documented in
three basins, the Central Andean Basin (Cordillera Oriental and Puna of Jujuy
and Salta provinces), the Precordillera Basin (La Rioja and San Juan provinces)
and the Famatina Basin (La Rioja province).
In
the Cordillera Oriental, the record of microflores ranges from the early
Tremadocian to the early Llanvirn, in the Famatina basin palynomorphs have a
middle Arenig age, while in the Puna and Precordillera, they are confined to the
Upper Ordovician (Puna: Ordovician-Silurian boundary; Precordillera: Hirnantian
and Llanvirn-Caradoc strata).
From
a paleogeographical point of view, Argentinian acritarch assemblages of the
Lower Ordovician belong to the Peri-gondwana Province (Tongiorgi et al.,
1994). Vavrdová (1974) defined the former Mediterranean Province, later
redefined and extended by different authors, and renamed as the Peri-Gondwana
Province. The Peri-Gondwana Province, as considered by Playford, Ribecai and
Tongiorgi (1995), comprises “a broad, latitudinally extensive, circumpolar,
cold to temperate paleogeographical belt along the northern Gondwana border,
from Argentina through eastern Newfoundland, north Africa, central and southern
Europe, and southern Turkey to South China”.
Afterwards,
the genera Arbusculidium, Coryphidium and Striatotheca have
been added as diagnostic elements of the Peri-Gondwana Province (Li Jun, 1987).
This province has also been identified in Sardinia (Albani, 1989), Pakistan
(Tongiorgi et al., 1994) and Iran (Ghavidel-Syooki, 1995).
Subsequently,
Servais and Fatka (1997) summarized the previously palaeobiogeographical
information from early to Middle Ordovician acritarchs. Likewise, high latitude,
cold to temperate water realm (climatically controlled) have been assigned to
Perigondwana assemblages, and low latitude, warm water realm to Baltica and the
East European Platform assemblages.
Recently
Rubinstein and Vaccari (2001) have reported acritarchs and cryptospores from the
upper siliciclastic member of the Salar del Rincón Formation. This unit crops
out in western flank of the Puna unconformably above Middle Ordovician volcanic
sequences. The sequence mainly bears brachiopoda and mollusca faunas attributed
to the latest Ashgill - earliest Llandovery (Benedetto and Sanchez, 1990).
Although the palynomorph assemblages from the upper member of the Salar del Rincón
Formation lacks enough markers to identify the stratigraphic Ordovician-Silurian
boundary, according to Rubinstein and Vaccari (2001), palynomorphs suggest a
latest Ordovician (post Hirnantian)-earliest Silurian (Rhuddanian) age,
supported by the presence of acritarchs as Eupoikilofusa striata and Dactylofusa
spinata (partial synonym of Dactylofusa estillis), and by the
occurrence of the genus Imperfectotriletes. The first occurrence of Imperfectotriletes,
as well as the absence of true trilete spores allow the correlation with the Imperfectotriletes
spp. Interval Biozone (sub-biozone a) of the Imperfectotriletes spp.-
Laevolancis divellomedia Assemblage Biozone of Steemans et al. (2000).
This zone is believed to correspond to the Hirnantian (uppermost Ordovician).
The difference between
the
faunal and palynological dates should be clarified with further study.
The
discovery of oldest cryptospores (probable bryophyte-like plants, sensu Gray,
1985; Wellman, 1999) can contribute much to evolutionary scenario of the early
land plants. The mention of them in Ordovician sequences of Argentina (Ottone et
al., 1999, Rubinstein, 2001, with references) similary to those found from
coeval deposits from around world (such as North Africa, the Prague Basin,
Sardinia, southwestern Wales, China, a.o. sensu Wang et al., 1997
and Wellman, 1999) suggests a cosmopolitan character of cryptospores, probably
confined to periodically damp environmental (Wellman, 1999)
Cordillera
Oriental
The Cordillera Oriental constitutes the southern part of the larger Central Andean Basin. The Santa Victoria Group represents the early Ordovician deposits. It includes the Santa Rosita Formation of a late Cambrian to late Tremadocian age, followed by the Acoite Formation (Lower to Middle
Figure 1.
Localities with Ordovician palinomorphs of Argentina (Puna, Cordillera Oriental, Famatina System and Precordillera basins).Arenig).
Both, Santa Rosita and Acoite Formations are composed of a thick sequence of
over 3.000 m dominantly clastic.
1.1.
Tremadocian
-
Yacones and La Caldera Region (San José Shales and Caldera Sandstone - La
Caldera department, Salta province). In the San José Shales and Caldera
Sandstone from the Yacones and La Caldera locality, eastern side of the
Cordillera Oriental, Bultynck and Martin (1982) presented the first discovery of
Tremadocian acritarchs in South America. The taxa mentioned in this region were Acanthodiacrodium
sp., A. angustum, Cymatiogalea cuvillieri, C. sp., Saharidia
sp., Vulcanisphaera sp. Although the authors mentioned the
impossibility to refer the taxa specifically to the Lower or Upper Tremadocian,
comparison with British, Spanish and French forms allowed them to assign a Lower
Tremadocian age. Associated invertebrate faunas confirm this age (e.g.
Harrington and Leanza, 1957).
-
Nazareno Region (Santa Victoria department, Salta province).
The microflore of this region (eastern slope of the Cordillera Oriental) has
been analyzed by Manca et al. (1995) and Rubinstein (1997). The
palynomorphs come from black shales of the lower part of the Santa Rosita
Formation and has been interpreted as transitional early to late Tremadocian age
from trilobite evidence. Complementary conodonts data from other horizons of
this formation indicated a late Tremadocian age (Manca et al. 1995). The
palynological sample yielded a poorly preserved acritarch assemblage with a
relatively high degree of thermal maturation. Despite their poor preservation
the following taxa have been identified: Acanthodiacrodium cf. simplex,
A. cf. spinum, A. tuberatum, A. spp., Actinotodissus achrasi, A. sp.,
Aremoricaniun sp., Cristalinium cambriense, Cymatiogalea cf bouvardii,
C. cristata, C. cf. multarea, C. cf. velifera, C. sp., Elektoriskos?
sp., Goniosphaeridium cf. akrochordum, G. pungens, Impluviculus cf.
lenticularis, I. Stellaris, Leiofusa spp., Leiosphaeridia spp., Micrhystridium
cf. nannacanthum, M. cf. robustum, M. shinetonense, Polygonium sp.,
Solisphaeridium sp., Stelliferidium cf. stelligerum,
Stelliferidium? sp., Synsphaeridium sp. and Vulcanisphaera cf.
frequens. According to Rubinstein (1997), the acritarchs clearly indicate
a Tremadocian age, although has not been possible to assign either to a Lower or
Upper Tremadocian. Paleogeographically, the assemblage showed affinities with
the Mediterranean Paleoprovince (Vavrdová 1974; Martin, 1982) and thus a high
paleolatitudinal location have been inferred to the Tremadocian Nazareno
assemblage.
Abra
de Zenta Region (Humahuaca department, Jujuy province). In the eastern flank of the Cordillera Oriental, within
the lower levels of Santa Rosita Formation cropping out at the Abra de Zenta, Aráoz
and Vergel (2001) and Aráoz (2002) described a marine plankton assemblage with
the following
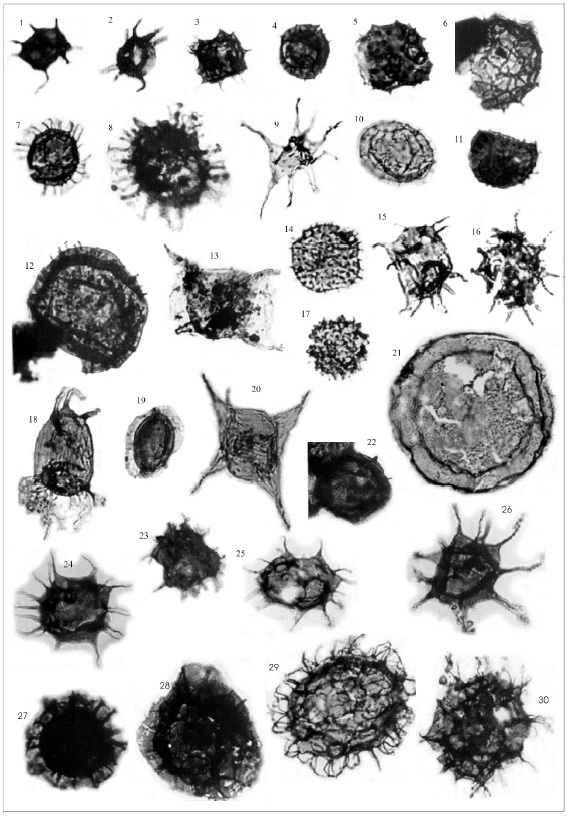
Plate I: 1-Micrhystridium shinetonense Downie, 1958. 2-Impluviculus stellaris Martin, 1977. 3-Vavrdovella areniga (Vavrdová) Loeblich y Tappan, 1976. 4-Stelliferidium? simplex (Deunff, 1961) Deunff et al., 1974. 5-Coryphidium sp. 6-Dictyotidium sp. 7-Athabascaella? sp. 8-Ordovicidium sp. cf. O. yangtzeense Tongiorgi et al., 1995. 9-Tectitheca sp. cf. T. additionalis Burmann, 1968. 10-Cymatiosphaera sp. 11-Stelliferidium sp. cf. S. brevipalmatum Albani, 1989. 12-Rhopaliophora sp. cf. R. pilata (Combaz y Peniguel, 1972) Playford y Martin, 1984. 13-Aureotesta clathrata var. simplex (Cramer et al., 1974) Brocke, 1997. 14-Coryphidium miladae - C. tadla group sensu Rubinstein et al., 1999. 15-Barakella sp. cf. B. rara (Lu, 1987) Tongiorgi et al., 1995. 16-Stellechinatum uncinatum (Downie, 1958) Molyneux, 1987. 17-Buedingiisphaeridium tremadocum Rasul, 1979. 18-Arbusculidium filamentosum (Vavrdova, 1965) Vavrdova 1972. 19-Dactylofusa velifera Cocchio forma brevis Albani, 1989. 20-Eisenackidium orientalis Rubinstein, 1999. 21-Saharidia fragilis (Downie 1958) Combaz 1967. 22-cf. Acanthodiacrodium angustum (Downie 1958) Combaz 1967. 23-Acanthodiacrodium tuberatum (Downie) Martin 1973. 24- Acanthdiacrodium crassus (Loeblich y Tappan 1978). 25-Acanthodiacrodium achrassi (Martin 1973) Yin Leiming, 1986 26- Polygonium gracile Vavrdova 1966. 27-Cymatiogalea velifera (Downie 1958) Martin 1968 28-Cristallinium cambriense (Slavikovà) Vanguestaine 1978. 29-Vulcanisphaera africana (Deunff 1961) Rasul 1976. 30-Vulcanisphaera turbata Martin 1981. (From: 1,2: Rubinstein, 1999 (not scale assigned). 3-20: Rubinstein et al., 1999 (3,4,7: 500x - 5,9,14-17,19: 375x - 6,8,10- 13,18,20: 250x). 21-30: Aráoz, 2002 (21,26,29,30: 250x - 22: 500x – 23-25,27,28: 375x).
taxa:
Acanthodiacrodium achrassi; A. crassus, A. formosum, A.
hirsutum, A. ignoratum, A. tuberatum, A. cf. angustum, Cristallinium
cambriense, Cymatiogalea velifera, C. sp., Leiosphaeridia sp., Lophosphaeridium
spp., Polygonium dentatum, P. gracile, P. symbolum, cf. P.
pungens, Polygonium spp., Saharidia downie, S. fragilis, Stelliferidium spp.,
Vulcanisphaera africana, V. nebulosa, V. tuberata and V. turbata.
The
microplankton assemblage is more diversified that others previously found in the
early Tremadocian strata of northwestern Argentina. However this assemblage was
considered as low to moderate diversity, dominated by Saharidia spp
(76,2%), leiosphaerids (9,1%), Polygonium (5,1%) and acanthomorphitae
forms (3,6%) as the less frequent. The assemblage was interpreted as indicative
of a restricted marine setting of a shallow-water environment with Saharidia and
leiosphaerids dominants (following Vecoli, 2000). The analyzed assemblage has
been referred to the early Tremadocian, but did not provide yet enough
diagnostic taxa to secure a full affinity with the Perigondwanian Province
(Playford et al., 1995).
1.2.
Upper Tremadocian – Arenig
-
Santa Victoria Area (Santa Victoria department, Salta province).
An integrated biostratigraphic scheme obtained from acritarchs, graptolites, and
trilobites of an Upper Tremadocian to Middle Arenig age has been recently
established in the Santa Victoria area belonging to eastern side of the
Cordillera Oriental of NW Argentina (Rubinstein et al., 1999; Rubinstein
and Toro, 1999; 2001).
Five
palynological assemblages have been recognized from the uppermost Tremadocian
(Santa Rosita Formation) and the Lower-Middle Arenig (Acoite Formation) at the
Rio La Huerta and Rio Acoite sections (Rubinstein et al., 1999). In
ascending order, the following biozones have been established:
Assemblage
1: From the Upper Tremadocian, equivalent to the Adelograptus-Bryograptus
graptolite association, is characterized by Cymatiogalea cristata, C.
velifera, C. cuvillieri and Vulcanisphaera sp.
Assemblage
2: From the Lower Arenig, equivalent to the Tetragraptus
akzharensis graptolite zone.
The
first appearance of the genus Coryphidium, and the following species has
been registered:
Dactylofusa
velifera forma brevis, Eisenackidium orientalis, Vavrdovella
areniga and Veryhachium sp. cf. rhomboidium. In addition, Rhopaliophora
palmata and R. sp. cf. pilata were restricted to this
assemblage in the Río Acoite section. Other species exclusive to this
assemblage were Acanthodiacrodium sp. cf. tasselli, Buedingiisphaeridium
tremadocum, Cymatiogalea sp. cf. membranispina, Micrhystridium
henryi, Ordovicidium sp. cf. yangtzeense, Stelliferidium sp.
cf. brevipalmatum and Vavrdovella areniga.
Assemblage
3: This assemblage has been recognized in the lower part
of the Baltograptus deflexus graptolite zone (Lower Arenig). The only
species to appear is Cymatiogalea granulata.
Assemblage
4: From the upper part of the B. deflexus graptolite
zone (Lower Arenig). It is characterized by the first appearance of the genus Striatotheca,
with the species S. cf. principalis var parva and Ammonidium
multipugiunculatum.
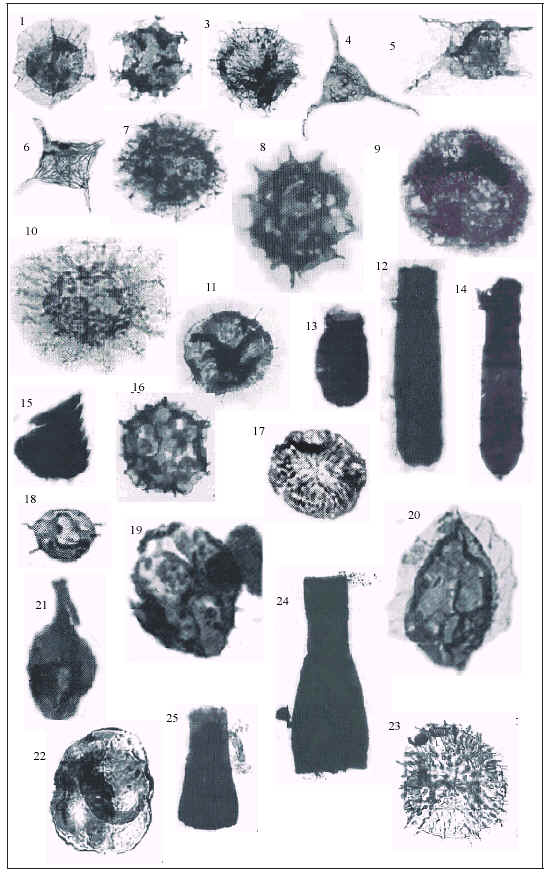
Plate II: 1-Cymatiogalea granulata Vavrdova, 1966. 2-Adorfia hoffmanensis (Cramer et al., 1974) Ottone, 1992. 3-Ammonidium ballistum Ottone, 1992. 4-Veryhachium trispinosum (Eisenack, 1938) Deunff, 1954. 5-Marrocanium simplex Cramer et al., 1974. 6-Striatotheca principalis Burmann, 1970. 7-Peteinosphaeridium trifurcatum intermedium Eisenack, 1976. 8-Vulcanisphaera tuberata (Downie 1958) Eisenack et al., 1973. 9-Asketopalla sp. 10-Multiplicisphaeridium multipungilatum Cramer y Diez, 1977 11-Baltisphaerosum peniguelii Ottone, 1992 12-Spathachitina sp. A. 13-Lagenochitina sp. A. 14-Conochitina sp. A. 15- Scolecodont sp. A. 16-Cristallinium chalalensis Ottone, 1995. 17-Virgatasporites sp. similar V. rudi Combaz, 1968. 18- Acanthodiacrodium cf. A. simplex Combaz, 1968. 19-cf. Baltisphaeridium hirsutoides hamatum Downie, 1958. 20-Pterospermella crameri (Volkheimer et al., 1980). 21-Pirea sp. cf. Pirea dubia Vavrdova, 1972. 22-Tetrahedraletes sp. 23-Gorgonisphaeridium granulobrachium Ottone, 1999. 24-Cyathochitina sp. cf. C. jenkinsi Neville, 1974. 25-Belonechitina sp. A. (From: 1-15: Ottone, 1992 (1-7,11,15: 250x - 8: 750x - 9: 375x - 10: 500x – 12-14: 75x). 16: Ottone et al., 1995 (x750). 17-19: Volkheimer et al., 1980 (17-19: 750x). 20-23: Ottone et al., 1999 (x375). 24, 25: Ottone et al., 2001 (26,27: 75x).
Assemblage
5: Assemblage recorded from the Didymograptellus
bifidus graptolite zone (Middle Arenig). It is characterized by the
increasing abundance and diversity of acritarchs and by the first occurrence of
the following taxa: Acanthodiacrodium costatum, Adorfia hoffmanensis,
Arbusculidium filamentosum, Aureotesta clathrata var. simplex,
and Coryphidium miladae-C. tadla group. Also, restricted to lower D.
bifidus graptolite zone, the following taxa appear: Barakella sp. cf.
rara, Coryphidium australe, Petaloferidium sp. cf. florigerum,
Polygonium sp. cf. delicatum, P sp. cf. symbolum, P.
spp., Striatotheca microrugulata, S. sp. cf. rarirrugulata,
S. transformata, and Tectitheca sp. cf. additionalis.
Recently,
Rubinstein and Toro (1999; 2001) strengthened the former biostratigraphical
scheme with two new sections (Quebrada Grande and Quebrada de La Curva), adding
new data from La Huerta section (Jujuy province). From the middle part of the
Quebrada Grande section Rubinstein and Toro (1999) noticed the first report of Cymatiogalea
messaoudensis var. messaoudensis. The occurrence of this species
which is diagnostic of the messaoudensis-trifidum Assemblage
(latest Tremadocian to earliest Arenig) allowed some considerations. The record
of Cymatiogalea messaoudensis var. messaoudensis is accompanied by
Arbusculidium filamentosum, Coryphidium cf. bohemicum, Dactylofusa
velifera forma brevis and Eisenackidium orientalis among
others. So the occurrence of Cymatiogalea messaoudensis var.messaoudensis
with Arbusculidium filamentosum suggests an age
similar to that of the sub-assemblage 5 of the messaoudensis-trifidum Assemblage
(base of the Arenig sensu Servais and Molyneux, 1997).
The
first appearance of Arbusculidium filamentosum has been reassigned as
corresponding to lowest B. defexus Zone (early Arenig age) equivalent to
the former Assemblage 3. On the other hand, Rubinstein and Toro (2001)
mentions Cymatiogalea messaoudensis var. messaoudensis from the
Middle Arenig (Didymograptellus bifidus Zone) of the La Huerta section,
suggesting that this species may range higher in Quebrada Grande section than
previously thought. Acritarch assemblages from Quebrada La Curva section have
been correlated with the Didymograptellus bifidus Zone and assigned to a
Middle Arenig age.
According
to Rubinstein and Toro (1999; 2001) some paleogeographical inferences can be
done on the basis of an integrated study with acritarchs, graptolites and
trilobites from Upper Tremadocian to the Middle Arenig in the Cordillera
Oriental of Argentina. Perigondwanan affinities have been interpreted for the
assemblages of NW Argentina, during the early Ordovician (early to middle
Arenig). The palynomorphs from the Acoite Formation indicated a temperate
paleolatitudinal location for northwestern Argentina, transitional, near the
limit of the cold water Perigondwana Realm (units climatically controlled as
suggested Servais and Fatka, 1997). Some elements are shared with those of
intermediate paleolatitudinal locations such as South China and Pakistan. These
data confirms the paleogeographical location of the Cordillera Oriental during
the early Ordovician, previously determined by trilobites and graptolites
(Waisfeld, 1993; Toro, 1996). An unrestricted communication between the eastern
and western side of the Cordillera Oriental have been confirmed and clarified
(Astini and Waisfeld, 1993; Waisfeld, 1996; Moya, 1988; Moya and Montero, 1999).
Both,
biostratigraphical and paleogeographical implications of Arenig palynomorphs
include data from Los Colorados Creek and Rio Cajas localities (Ottone et al.,
1992) to the western side of the Cordillera Oriental.
-
Parcha Region (La Caldera department, Salta province). In the southern part of the
Cordillera Oriental, the Parcha section has been palynologicaly analyzed by
Rubinstein at al. (1999) and Rubinstein and Toro (1999). Acritarchs were
recovered from the Saladillo Formation (partially equivalent to the Santa Rosita
Formation) and Parcha Formation (partially equivalent to the Acoite Formation).
The palynofloras are poor in abundance, diversity, and preservation. Thus ages
and correlations have been based on the graptolites. Samples from the lower part
of the succession corresponding to Saladillo Formation (Tremadocian) yielded Bryograptus
sp. and the following palynomorphs:
Polygonium
dentatum and Baltisphaeridium cf. multipilosum. In
the Parcha Formation (Lower Arenig at Parcha section) the basal sample
equivalent to the Tetragraptus akzharensis graptolite zone has yielded Stelliferidium
sp. and Actinotodissus? sp. The following Baltograptus deflexus graptolite
zone contain Stellechinatum cf. uncinatum, Micrhystridium
stellatum, Peteinosphaeridium sp., Cymatiogalea sp., Cymatiosphaera
sp., Coryphidium sp., Leiosphaeridium sp., Acanthodiacrodium
spp. and Polygonium spp. Recent acritarch findings in Parcha Region
(Rubinstein and Toro, 2002) lead to support the presence of the messaoudensis-trifidum
acritarch assemblage in the Tremadocian-Arenigian boundary of northwestern
Argentina. Recorded species are Caldariola glabra var. glabra, Cymatiogalea
messaoudensis var. messaoudensis, Cymatiogalea messaoudensis var.
inconnexa, Cymatiogalea deunffi, Coryphidium sp., Stelliferidium
cf. trifidum and Acanthodiacrodium-Vavrdovella transients.
Acritarchs come from a level related to the Araneograptus murrayi graptolite
biozone, which indicates an age not younger than late Tremadocian for the
acritarch assemblage.
-
Sierra de Cajas Region and Los Colorados Area (Humahuaca and Tumbaya department,
Jujuy province).
The
palynomorphs studied by Ottone et al. (1992) in this region (western
slope of the Eastern Cordillera) have been recovered from the Acoite Formation.
It is a clastic sequence mainly composed of thinly laminated claystones,
siltstones and sandstones. This unit rests conformably above the Tremadocian
Santa Rosita Formation and the Upper Arenig-Lower Llanvirn Sepulturas Formation
conformably overlies it. The palynological samples are from the lower and upper
section of Acoite Formation at Los Colorados Creek (Tumbaya department), and
from the lower section of the unit at the Rio Cajas (Humahuaca department). Two
distinct palynomorph assemblages with acritarchs, algae, chitinozoans,
scolecodonts and graptolite siculae have been differentiated throughout the
unit: a lower Acoite Palynological Assemblage (AL) and an Upper Acoite
Palynological Assemblage (AU). The following taxa are present: Acanthodiacrodium
cf. angustizonale, A. spp., Adorfia hoffmanensis, Ammonidium
ballistum, Arbusculidium sp., Asketopalla sp., Baltisphaerosum
peniguelli, Cristallinium sp., Cymatiogalea granulata, C. velifera,
Eisenackidium sp., ?Gloecapsomorpha sp., Leiosphaeridia sp., Lophosphaeridium
aequicuspidatum, Marrocanium simplex, Micrhystridium henryi,
Multiplicisphaeridium multipungilatum, M. spp., Peteinosphaeridium
trifurcatum intermedium, Polygonium gracile, Striatotheca principalis, S.
transformata, S. triangulata, Veryhachium lairdii, V. trispinosum,
Vulcanisphaera tuberata, Conochitina sp., Lagenochitina sp., Spatachitina
spp., Cyathochitina sp. and Scolecodont sp. The AL Assemblage has
been characterized by a relative scarcity of palynomorphs and a poor
preservation. The most abundant and consistently represented form is Leiosphaeridia
sp. (ca.50%); the minor constituents of the assemblage are Vulcanisphaera
tuberata (ca. 5,5%), Cymatiogalea velifera (ca. 5%), and Polygonium
gracile (ca. 5%). Graptolite siculae are relatively common and the Thermal
Alteration Index (TAI) (Staplin, 1969) varies from 3 (rarely) to more than 4. An
early Arenig age was referred to the AL Assemblage from the lower section of the
Acoite Formation, equivalent to upper part of the Baltograptus defexus graptolite
zone (Rubinstein and Toro, 2001).
The
AU Assemblage has been characterized as moderately diverse and relatively
well-preserved palynological assemblage. The commonest taxon is Polygonium
gracile (ca. 33%), together with a consistent representation of Cymatiogalea
granulata (17%), leiosphaerids (10%), Veryhachium lairdii (7%) and Adorfia
hoffmanensis (5%). The TAI assigned to thinner-walled acritarchs was of
about 2 +.
The
AU Assemblage from the upper section of the Acoite Formation has been referred
to Middle Arenig, equivalent to Didymograptellus bifidus graptolite zone
(sensu Rubinstein and Toro, 2001).
Both
AL and AU palynological assemblages have been related to the Mediterranean or
Perigondwana Province (Vavrdová, 1974, Martin, 1982; Li Jun, 1987; Albani,
1989) with an inferred high latitudinal setting.
-
Chalala Creek (Tumbaya department, Jujuy province).
Also on the western side of the Cordillera Oriental from the Chalala Creek, near
Purmamarca locality, Ottone et al. (1995) described a scarce a relatively
low-diversified acritarch assemblage from the basal section of the Acoite
Formation. The following taxa have been identified: Acanthodiacrodium spp.,
Baltisphaerosum sp., Cristallinium chalalensis, Cymatiogalea velifera,
Leiosphaeridia sp., Lophosphaeridium aequicuspidatum, Polygonium gracile,
Vulcanisphaera tuberata, and undetermined chitinozoans and scolecodonts.
This
assemblage showed some resemblance with the previously defined AL Assemblage
from the Acoite Formation at Rio Cajas and Los Colorados sections, which has
been referred to early Arenig (Ottone et al., 1992). However, owing to
the absence of Striatotheca and conspicuous forms of the AL Assemblage
such as: Peteinosphaeridium trifurcatum intermedium and Baltisphaerosum
peniguelli the authors suggested a probably Tremadocian age to the
assemblage, but not similar to the previously known assemblage from Tremadocian
strata of the Cordillera Oriental (Bultynck and Martin, 1982).
This
microflore have also been included in the Mediterranean or Perigondwanian
Province 1.3. Llanvirn
- Mojotoro Region (Quebrada del Gallinato and Corral de Barranca, Sierra de
Mojotoro, Capital department, Salta province). The
first Ordovician acritarchs mentioned in South America were localized in this
region. In the upper part of the Mojotoro Formation two palynoassemblages have
been documented by Volkheimer et al. (1980b). Both assemblages belong to
an only microfloristic unit and the palynomorphs (acritarchs and prasinophytes)
recorded are Acanthodiacrodium cf. latizonale, A. cf. simplex,
Baltisphaeridium cf. redondensis, cf. Baltisphaeridium hirsutoides
hamatum, Cymatiosphaera spp., Leiovalia sp., Leiosphaeridia spp.,
Multiplicisphaeridium cf. varians, Pirea cf. dubia, P. sp. Pterospermella
crameri, Veryhachium sp. and Virgatasporites cf. rudi,
and chitinozoans (gen. et sp. indet.).
According
to Volkheimer et al. (1980b) the stratigraphic value of palynomorphs
indicated a Lower Ordovician to Llanvirn age for the Mojotoro Formation. In
addition, it was correlated to other Lower Ordovician assemblages from the
Sahara. A narrow paleogeographical relationship between both, northwest
Argentina and North Africa, have been inferred for the early Ordovician.
2.-
Precordillera Basin
The
Precordillera of western Argentina has been a matter of intense debate during
the last decade. This debate is focused on its supposed paraautochthonous –
allochtonous origin. These problematic are suggested on the basis of tectonic,
stratigraphic and paleontological basis (Bond et al., 1984; Ramos, 1986;
1988; Dalla Salda et al., 1992; Dalziel et al., 1996; Astini et
al., 1996; Pankhurst and Rapela, 1998; Benedetto et al., 1999; Aceñolaza
and Toselli, 1999; Finney et al., in press, with references).
Ottone
et al. (1999) and Ottone et al. (2001) interpretates a Gondwanan
link during the Llanvirn for the supposed allochtonous Precordilleran terrane on
the basins of palynomorphs Carbonate and siliciclastic rocks ranging in age from
the Tremadocian to the Ashgill mainly characterize the Ordovician sedimentary
sequences from Precordillera Basin. An extensive carbonate platform with thick
limestone sequences (San Juan Formation) was established during the Cambrian and
early Ordovician for the Eastern and Central Precordillera. In the western part
of the basin, the carbonate sedimentation persisted until the early Caradoc (Las
Aguaditas Formation). Late Tremadocian and Arenig alternating clastic
sandstones, mudstones, and black shales assigned to the Gualcamayo and Los
Azules formations were deposited in several places of Precordillera; these
sequences were related to an Arenig-Llanvirn transgresive event. The following
Middle and Upper Ordovician successions include shales, sandstones, and
conglomerates (Las Plantas, Trapiche, Don Braulio, and La Chilca formations)
were deposited in several parts of the basin until the uppermost Ashgill.
The
Ordovician sequences display abundant fossil faunas (graptolites, conodonts,
trilobites, brachiopods, a.o.) that have been treated in several papers and
provided valuable data to understand the regional paleoenvironmental evolution.
For the present, palynomorphs have only been recovered from early Ordovician
levels (San Juan, Gualcamayo and Los Azules Formations) and for the Hirnantian,
uppermost Ashgill (La Chilca Formation).
2.1.
Arenig/Llanvirn
-
Jachal Region (Jachal department, San Juan province).
Enigmatic organic-walled microfossils have been documented from two sections
near Jachal locality (Heuse et al., 1997). The incertae sedis microfossils
from Cerro La Silla section come from lower levels of San Juan Formation (early
Arenig) correlated with upper part of the P. elegans / O. communis conodont
zone. The other section outcropping at the Cerro La Chilca yielded material from
early Llanvirn strata correlated with E. suecicus conodont zone from the
top of the San Juan Formation and the basal part of the Gualcamayo Formation.
These problematic microfossils were described as black, spherical to
subspherical shaped forms with an opening and cell-like wall structure. The
possible biological affinities was discussed by Heuse et al. (1997) and a
hypothetic origin, linked to chaetognath specimens have been assigned from
similar Bohemian microfossils founded inside two specimens of the chaetognath Titerina
rokycanensis.
-
Guandacol Region (Quebrada Potrerillos, General Lavalle department, La Rioja
province). The
first data on Ordovician chitinozoans from Argentina belong to Volkheimer (1978)
and Volkheimer et al. (1980a). The chitinozoans have been recovered from
dark shales of the Gualcamayo Formation (early Llanvirn) at Quebrada
Potrerillos, southwest of Guandacol locality. The assemblage has been
characterized by dominant big sized forms (130-470 µm) with smooth or not much
ornamented vesicle wall and a simple carina. Not appendices or siphoned forms
have been observed. It belongs to the genera Lagenochitina, Conochitina,
Cyathochitina, and Rhabdochitina? and Desmochitina.
Considerations
about evolutionary successions on carina and appendices presence have been
analyzed to establish an approximate age. Also the stratigraphic value of
individuals forms such as Cyathochitina and Conochitina and its
related species allowed to assign for the analyzed levels a Llanvirn age. The
Gualcamayo Formation has been mainly dated as of an early Llanvirn age by
graptolite record (Cuerda and Alfaro, 1986).
Equivalent
levels on the west flank of Cerro Perico at the Potrerillos creek have yielded
scolecodonts (Ottone and Holfeltz, 1992). The scolecodonts or fossilized
elements of the proboscidal armatures of polychaetous annelids that occur in
many types of marine sediments. They are abundant in shallow water and appear
often associated with graptolites, chitinozoans, spores and acritarchs. The
oldest scolecodonts have been found in Arenig rocks. To the Gualcamayo Formation
the following taxa have been recognized: Anisoceracites sp., Arbellites
sp., Leodicites sp., Nereidavus sp., Nereigenys sp. and
Staurocephalites sp. The early Llanvirn age assigned to the assemblage
have been obtained by the associated marine biota.
2.2.
Llanvirn/Caradoc
-
Cerro Viejo de Huaco Region (Jachal department, San Juan province). Middle
Ordovician sequences belonging to Los Azules Formation crops out in the Cerro
Viejo de Huaco region, between Huaco and Jachal localities, northern San Juan
province, in the Central Precordillera. Los Azules Formation is mainly made up
of argillites, shales and mudstones, and was divided into three informal members
(Ortega, 1987). From these levels Ottone et al. (1999) reported a rich
association of micro and macrofossils with palynomorphs, conodonts and
graptolites associates. Several species of acritarch (Baltisphaerosum
peniguelli, B. spp., Gorgonisphaeridium granulobrachium, Lophosphaeridium
ruidum, Navifusa similes, Ordovicium elegantulum, Orthosphaeridium vibrissiferum,
Polygonium echinatus and Solisphaeridium sp.), prasinophytes (Leiosphaeridia
sp., Pterospermella sp.), cryptospores (Tetrahedraletes sp.)
and different morphotypes of phytodebris were identified throughout the middle
and upper members of the Los Azules Formation. Several species of conodonts have
also been recorded within the middle member of the unit.
Considering
the stratigraphic value and occurrence of each species in Los Azules section,
two assemblages were defined: the Lower Los Azules Palynological Assemblage
(LAL), and the Upper
Los
Azules Palynological Assemblage (LAU). The first spans the entire middle member
of the unit.
Leiosphaerids
are the commonest forms (83-98%); Navifusa similis and smooth
chitinozoans are subordinate. Terrestrial remain have not been observed. The
second, in the upper member of the formation, yielded plentiful ornamented
chitinozoans (60-81%); also Navifusa similis (1-10%); Ordovicidium
elegantulum (1-7%), Leiosphaeridia sp. (2-5%), scolecodonts (2-5%)
and Lophosphaeridium ruidum (3-4%) as the minor constituents. Terrestrial
palynomorphs are rare (less than 1%).
According
to Ottone et al (1999) paleoenvironmental inferences related to low
diversity, leiosphaerids dominated assemblages are indicatives of restricted
epicontinental deep-water settings to the middle member of the Los Azules
Formation. In addition, the assemblage of the upper member of the unit is
indicative as well as, to an offshore setting, in agreement with the
sedimentological and tectonic data (Astini, 1994a,b; 1995; Baldis et al., 1989).
The
LAL Assemblage lacks enough plant microfossils markers to confirm a precise
stratigraphic rank, and was attributed to a late Llanvirn age on the basins of
associated fauna. The presence of chitinozoans was later analyzed by Ottone et
al. (2001) confirming this late Llanvirn age. On the other hand, the LAU
Assemblage was biostratigraphically placed in the Llanvirn/Caradoc, although the
chitinozoan assemblages suggested a younger age (early Caradoc). Chitinozoans
species of the LAL Assemblage (middle member of the Los Azules Formation) are: Calpichitina
megastrophica, C. sp. A; Cyathochitina dispar, C. sp.
cf. campanulaeformis, C. sp. cf. jenkinsi, Conochitina dolosa,
C. minnesotensis, Desmochitina minor forma amphorae, Desmochitina
minor forma ovulum, Desmochitina minor forma typical, Hyalochitina
sp., Lagenochitina cilindrica and L. sp. cf. baltica.
Also the following chitinozoans have been reported from the upper member of Los
Azules Formation or in the LAU Assemblage: Angochitina sp., Armoricochitina
sp. cf. nigerica, Belonechitina punctata, B. sp., Colpichitina
sp., Conochitina dolosa, C. minnesotensis, Conochitina?
pygmaea, Cyathochitina sp. cf. campanulaeformis, C.
sp. cf. jenkinsi, Eisenackitina yolei, Hercochitina
volkheimerii, Kalochitina sp. cf. multispinata.
All
palynomorph assemblages confirm the previous graptolite-based age for Los Azules
Formation (early Llanvirn-early Caradoc) and exhibits similarities with coeval
assemblages from the Northern Hemisphere and Gondwanaland. However the
appearance of some taxa such as Conochitina? pygmaea and Armoricochitina
sp. cf. nigerica in younger levels than in Laurentia and Northern
Gondwana was interpreted as an earlier radiation originated in Precordillera
with posterior migration and reaching their acme in Laurentia and Northern
Gondwana.
2.3.
Hirnantian
-
Talacasto area (Ullúm department, San Juan Province). In this area three
palynomorph assemblages have been described for the La Chilca Formation (late
Hirnantian-Llandovery/Wenlock?) and the Los Espejos Formation (Wenlock to
Pridoli) (Rubinstein and Brussa, 1999). The Assemblage 1 corresponds to the Normalograptus
persculptus graptolite zone of Hirnantian (latest Ordovician) age. It is
dominated by simple spheroidal and short-spined acritarch taxa, none of which
have any stratigraphic value. Genera include Leiosphaeridia, Michrystridium, and
Veryhachium.
3.-
Famatina Basin
The
Ordovician sedimentary rocks of the Famatina System are represented by isolated
outcrops between widespread intrusives. The sedimentary sequences belong to
clastic rocks (Upper Cambrian- Tremadocian) from the Central Famatina and
volcaniclastic rocks (Arenig) from Northern and Central Famatina. The
volcaniclastic sequences were included in the Famatina Group (Turner 1964) that
is integrated by the Suri (early to middle Arenig) and Los Molles (middle to
late Arenig) Formations. Later Aceñolaza and Toselli (1981) proposed the
Cachiyuyo Group to unify the clastic and volcaniclastic rocks. The latter is
integrated in ascending order by the following formational units: The Volcancito
(recently assigned to uppermost Cambrian to Tremadocian), Portezuelo de las
Minitas (= La Alumbrera, assigned to earliest Arenig), Suri (early to middle
Arenig), Los Molles (middle to late Arenig) and Morado (= Las Planchadas,
Llanvirn) formations. The sequence represent a continuous sedimentation during
the early Ordovician, controlled by volcanism, tectonic and eustatic changes.
The age assigned to the formational units have been extracted from unified
paleontological and biostratigraphical data of a recent synthesis (In Aceñolaza
et al., 1996; Astini, 1999 and Esteban et al., 1999 among others).
3.1.
Arenig
-
Saladillo Grande and Los Molles Creeks (Famatina department, La Rioja province).
Strata belonging to the Suri Formation cropping out at the Quebrada Saladillo
Grande and the sequences of Los Molles Formation from the Quebrada Los Molles
have yielded acritarchs and chitinozoans (Rubinstein and Astini, 2000 and
Rubinstein, 2001a,b). Acritarch assemblages from the Suri Formation have a
middle Arenig age, independently dated by conodonts (upper part of the Oepikodus
evae Zone).
Graptolites
of the Didymograptellus bifidus Zone have also been recovered in this
unit, bellow levels with acritarchs. This formation contains, among other
acritarch taxa Arbusculidium filamentosum, Eisenackidium orientalis,
Dactylofusa velifera forma brevis, Cymtiogalea deunffi, Vavrdovella
areniga, Striatotheca sp., Acanthodiacriodium costatum, Rhopaliophora
palmata, Poikilofusa striatogranulata and Peteinosphaeridium spp.
The chitinozoans are actually in study. The abundance of Eisenackidium
orientalis and Dactylofusa velifera forma brevis of an
outstanding stratigraphic value in the Cordillera Oriental (early to middle
Arenig), allowed a biostratigraphic correlation of both basins (Acritarch Assemblage
4 to 5 or Baltograptus deflexus-Didymograptellus bifidus graptolite
zones of the Cordillera Oriental). Thereby, the Suri Formation showed clear
perigondwanan affinities (moderate - cold water) with typical forms such us Arbusculidium,
Striatotheca, and taxa from temperate to low paleolatitudinal location.
All
typical perigondwanan acritarchs disappear towards the upper Los Molles
assemblages.
These
are replaced by intermediate to low latitudes taxa such as Chinese, Australian
and Baltic forms with predominance of acanthomorphic elements (Peteinosphaeridium,
Baltisphaerosum and Tongzia).
They
are also included in levels of the upper part of the Oepikodus evae Zone,
consequently corresponding to the Middle Arenig. According to Rubinstein (2001a)
the differences between Suri and Los Molles acritarch assemblages were due to
local environmental conditions related to the volcanic arc setting.
Aknowledgements. We are gratefull to G. Aceñolaza for valuable discussion on the stratigraphy of NW Argentina. We also thank the editor F. Aceñolaza, for the invitation to participate in the volume.
References
Aceñolaza,
F.G., Miller, H. and Toselli, A.J. 1996. Geología del Sistema de Famatina. Münchner
Geologische Hefte, A19. 410 pp.
Aceñolaza,
F.G. and Toselli, A.J. 1981. Geología del Noroeste Argentino. Facultad de Ciencias Naturales, Universidad
Nacional de Tucumán. 212 pp.
Aceñolaza,
F.G. and Toselli, A.J. 1999. Argentine Precordillera: Allochthonous or
Autochthonous Gondwanic? Zentralblatt fur Geologie und Palaeontologie.
Teil 1, Heft 7-8: 1-14.
Albani,
R. 1989. Ordovician (Arenig) Acritarchs from the Solanas Sandstone Formation,
Central Sardinia, Italy. Bolletino della Societá Paleontologica Italliana. 28
(1): 3-37.
Aráoz,
L. 2002. Palinología de los niveles inferiores de la Formación Santa
Rosita, en el Abra de Zenta, provincias de Salta y Jujuy, República Argentina.
Seminario Inédito. Facultad de Ciencias Naturales e I.M.L, Universidad Nacional
de Tucumán. 187
pp.
Aráoz,
L. and Vergel, M. d. M. 2001. Acritarcos del Ordovícico Inferior (Formación Santa Rosita) en el
Abra de Zenta, Cordillera Oriental, Argentina. V Jornadas de Comunicaciones. Serie
Monográfica y Didáctica. Nº 41. Facultad de Ciencias Naturales e I.M.L,
Universidad Nacional de Tucumán. p. 55.
Astini,
R. A. 1994a. Geología e interpretación de la Formación Gualcamayo en su
localidad clásica (suroeste de Guandacol y cordón de Perico-Potrerillo),
Precordillera septentrional. Revista de la Asociación Geológica Argentina.
49 (1-2): 55-70.
Astini,
R. 1994b. Análisis secuencial y paleoambientes de las pelitas negras
(aloformación Gualcamayo) que suprayacen a las sucesiones carbonáticas
eo-ordovícicas en la Precordillera argentina. Revista de la Asociación Geológica
Argentina. 49 (1-2): 71-84.
Astini,
R. A. 1995. Sedimentología de la Formación Las Aguaditas (talud carbonático)
y evolución de la cuenca precordillerana durante el Ordovícico medio. Revista
de la Asociación Geológica Argentina. 50 (1-4): 143-164.
Astini,
R. A. 1999. El Ordovícico del Sistema del Famatina. In: Geología del
Noroeste Argentino. González Bonorino G., Omarini, R. and Viramonte, J. (eds.).
Relatorio XIV Gongreso Geológico Argentino. 1: 152-159.
Astini,
R. A. and Waisfeld, B.G. 1993. Análisis estratigráfico y paleoambiental del Ordovícico inferior
(Formaciones Acoite y Sepulturas) al Oeste de Purmamarca, Cordillera Oriental
jujeña. XII Congreso Geológico Argentino y II Congreso de Exploración de
Hidrocarburos. 1: 96-106.
Astini,
R. A., Ramos, V.A., Benedetto, J.L., Vaccari, N.E and Cañas, F.L. 1996 La
Precordillera: un terreno exóticoma Gondwana. XIII Congreso Geológico
Argentino y III Congreso de Exploración de Hidrocarburos. Actas. 5:
293-324.
Baldis,
B.A., Bordonaro, O, Armella, C., Beresi, M, Cabaleri, N., Peralta, S. and
Bastias, H. 1989. La cuenca paleozoica inferior de la Precordillera Argentina.
In: Cuencas Sedimentarias Argentinas. Chebli, G.A. and Spalletti, L.A.
(eds.). Serie
Correlación geológica 6. 101-121 pp.
Benedetto,
J.L. and Sánchez, T.M. 1990. Fauna y edad del estratotipo de la Formación Salar del Rincón
(Eopaleozoico, Puna, Argentina). Ameghiniana 27 (3-4): 317-326.
Benedetto,
J.L., Sánchez, T.M., Carrera, M. Brussa, E.D. and Salas, M.J. 1999.
Paleontological contraints on succesive paleogeographic positions of
Precordillera terrane during the early Paleozoic. In: Laurenti-Gondwana
correlations before Pangea. Ramos, V.A. and Keppie, J.D. (eds.). Geological
Society of America, Special Paper. 336: 21-42.
Bond,
G.C., Nickeson, P.A. and Kominz, M.A. 1984. Breakup of a supercontinent between
625 Ma and 550 Ma: new evidence and implitations for continental histories. Earth
and Planetary Sciences Letters. 70: 325-345.
Bultynck,
P. and Martin, F. 1982. Conodontes
et Acritarches de l’ Ordovicien Inférieur et acritarches du Silurien inférieur
de la partie septentrionale de la Cordillère Argentine. Bulletin de l’
Institut royal des Sciences naturelles de Belgique, Sciences de la Terre. 53
(4): 1-21.
Cuerda,
A. J. and Alfaro, M., 1986. Las graptofaunas de la Formación Gualcamayo en su
localidad tipo, Precordillera de La Rioja. IV Congreso Argentino de
Paleontología y Bioestratigrafía, Actas 1: 31-43.
Dalla
Salda, L.H., Cingolani, C. And Varela, R. 1992. The early Paleozoic orogeny belt
of the Andes in southern South America: result of Laurentia-Gondwana collision?.
Geology. 20: 617-620.
Dalziel,
IW.D., Dalla Salda, L.H., Cingolani, C. And Palmer, P. 1996. The Argentine
Precordillera: a Laurentian Terrane? Penrose Conference Report, GSA Today. 16-18.
Dean,
W.T. and Martin, F. 1992. Ordovician biostratigraphic correlation in southern
Turkey. In: Global Perspectives on Ordovician Geology. Webby, B.D. and
Laurie, J.R. (eds.). Proceedings of the 6th
International
Symposium on the Ordovician System. University
of Sydney, Australia. 195-203.
Esteban,
S., Tortello, F., Mángano, M.G., Buatois, L. and Aceñolaza, F.G. 1999.
Bioestratigrafía del Paleozoico Inferior del Sistema del Famatina. In: Geología
del Noroeste Argentino. González Bonorino G., Omarini, R. and Viramonte, J.
(eds.). Relatorio XIV Congreso Geológico Argentino. 1:
217-223.
Finney,
S.; Gleason, J., Gehrels, G; Peralta, S. and Aceñolaza, G. in press.
Early Gondwanan Connection for the Argentine Precordillera Terrane. Earth and
Planetary Sciences Letters.
Ghavidel-Syooki,
M. 1995. Palynostratigraphy and palaeogeography of a Palaeozoic sequence
in the Hassanakdar area, Central Alborz Range, northen Iran. Review of
Palaeobotany and Palinology. 86: 91-109.
Gray,
J. 1985. The microfossil record of early land plants: advances in understanding
of early terrestrialization, 1970-1984. Philosophical Transactions of the
Royal Society of London. 309B; 167-195.
Harrington,
H.J. and Leanza, A.F.; 1957. Ordovician trilobites of Argentina. University
of Kansas, Special Publications 1: 259 pp.
Heuse,
T., Lehnert, O. and Kraft, P., 1997. Organic-walled microfossils Insertae
sedis from the Ordovician of Argentine Precordillera and Bohemia. Acta
Universitatis Carolinae (Geologica). 40 (3-4): 425-439.
Li
Jun, 1987. Ordovician acritarchs from the Meitian Formation of Guizou
province, south-west China. Palaeontology. 30 (3): 613-634.
Manca,
N., Heredia, S., Hunicken, M. and Rubinstein, C. 1995. Macrofauna,
conodontes y acritarcos de la Formación Santa Rosita (Tremadociano), Nazareno,
provincia de Salta, Argentina. Boletín de la Academia Nacional de Ciencias
de Córdoba. 60
(3-4): 267-275.
Martin,
F. 1982. Some aspects of late Cambrian and early Ordovician acritarchs. In: Basset
M.G. and Dean, W.T. (eds.), The Cambrian - Ordovician boundary: sections, fossil
distributions, and correlations. National Museum of Wales, Geological Series
3: 29-40.
Moya,
M.C. 1988. Lower Ordovician in the Southern part of the Argentine Eastern
Cordillera. In: Bahlburg, H., Breitkreuz, CH. and Giese, P. (eds.). The
Southern Central Andes, Lecture Notes in Earth Sciences, 17: 55-69.
Moya,
M.C. and Monteros, J.A. 1999. El Ordovícico Tardío y el Silúrico en el borde occidental de la
Cordillera Oriental Argentina. XIV Congreso Geológico Argentino. 1:
401-404.
Ortega,
G. 1987. Las graptofaunas y los conodontes de la Formación Los Azules, Cerro
Viejo, zona de Huaco, departamento Jáchal, San Juan. Tesis Doctoral Inédita.
Facultad de Ciencias Exactas, Físicas y Naturales. Universidad Nacional de
Córdoba. 210
pp.
Ottone,
E.G.,Albanesi, G.L., Ortega, G. and Holfeltz, G.D., 1999. Palynomorphs,
conodonts and associated graptolites from the Ordovician Los Azules formation,
Central Precordillera, Argentina. Micropaleontology, 45 (3): 225-250.
Ottone,
E.G. and Holfeltz, G.D., 1992. Hallazgo de escolecodontes en la Formación
Gualcamayo, Llanvirniano Inferior, Argentina. VII Simposio Argentino de
Paleobotánica y Palinología. Asociación Paleontológica Argentina,
Publicación Especial Nº 2: 85-88.
Ottone,
E.G., Holfeltz, G.D, Albanesi, G.L. and Ortega, G. 2001. Chitinozoans from the
Ordovician Los Azules Formation,
Central Precordillera, Argentina. Micropaleontology. 47
(2): 97-110.
Ottone,
J.C., Toro, B.A. and Waisfeld, B.G. 1992. Lower ordovician palynomorphs from the
Acoite formation, Northwestern Argentina. Palynology. 16: 93-116.
Ottone,
E.G., Waisfeld, B.G. and Astini, R.A.. 1995. Acritarcas del Ordovícico Temprano de la Quebrada
de Chalala, Noroeste de Argentina. Ameghiniana 32
(3): 237-242.
Pankhurst,
R.J. and Rapela, C.W. 1998. The Proto-Andean Margin of Gondwana. The
Geological Society, London, Special Publication. 142: 384.
Playford,
G., Ribecai, C. and Tongiorgi, M. 1995. Ordovician acritarch genera Peteinosphaeridium,
Liliosphaeridium, and Cycloposphaeridium: morphology, taxonomy,
biostratigraphy, and palaeogeographic significance. Bolletino
della Societá Paleontologica Italiana. 34 (1): 3-54.
Ramos,
V.A. 1986. El diastrofismo oclóyico: un ejemplo de tectónica de colisión
durante el Eopaleozoico en el Noroeste Argentino. Revista del Instituto de
Geología y Minería-UNJu. 6: 13-28.
Ramos,
V.A. 1988. Late
Proterozoic-Early Paleozoic of South America: a collisional history. Episodes.
11 (3): 168-175. Rubinstein, C.V. 1997. Tremadocian acritarchs from
northwestern Argentina. Review of Palaeobotany and Palynology. 98: 41-45.
Rubinstein,
C.V. 2001a. Correlaciones
entre las cuencas ordovícicas y Silúricas de la Argentina basadas en el
estudio de acritarcos, criptoesporas y mioesporas. In: Correlaçao de
Seqüencias Paleozóicas Sul-Americanas. Melo,
J.H.G. and Terra, G.J.S. (eds.). Ciencia-Técnica-Petróleo. Seçao:
Exploraçao de Petróleo. 20: 19-24.
Rubinstein,
C.V. 2001b. Bioestratigraphic and palaeogeographic implications of Lower
Ordovician Acritarchs from the Eastern Cordillera and Famatina, northwestern
Argentina. Early Palaeozoic Palaeogeography and Palaeobiogeography of Western
Europe and North Africa. Université des Sciences et Technologies de Lille
(USTL), Lille. Abstracts: 57.
Rubinstein,
C.V. and Astini, R.. 2000.
Primer registro de palinomorfos arenigianos en las formaciones Suri y Molles,
región del Famatina, provincia de La Rioja, Argentina. XI Simposio Argentino
de Paleobotánica y Palinología. Resúmenes. pp. 98.
Rubinstein,
C. V. and Brussa, E. D. 1999. A palynomorph and graptolite biostratigraphy of
the Central Precordillera Silurian basin, Argentina. En : Tongiorgi, M. and
Playford, G. (eds.), 1999, Studies in Palaeozoic Palynology,
Selected
papers from the CIMP Symposium at Pisa, 1998: Bolletino della Società
Paleontologica Italiana, 38 (2-3): 257- 266.
Rubinstein,
C.V. and Toro, B.A. 1999. Acritarch and graptolite biostratigraphy in the lower
Arenig of the peri- Gondwana related Eastern Cordillera. Acta
Universitatis Carolinae (Geologica).
43
(1-2): 255-258.
Rubinstein,
C.V. and Toro, B.A. 2001. Review of acritarch biostratigraphy in the Arenig of
Eastern Cordillera, northwestern Argentina. New data and calibration with the
graptolite zonation. In: Contributions to Geology and Palaeontology of
Gondwana – In honour of Helmut Wopfner. Weiss R.H. (eds.). Geological
Institute, University of Cologne Germany. pp. 421-439.
Rubinstein,
C. and Toro, B., 2002 The messaoudensis-trifidum acritarch assemblage
(late Tremadoc-early Arenig) from Parcha, Eastern Cordillera, northwestern
Argentina. CIMP International Meeting and Workshops, Lille, France.
Palaeozoic
Palynology in the Third Millenium: new directions in acritarch, chitinozoan and
miospore research. Abstract Volume, p. 48.
Rubinstein,
C.V., Toro, B.A. and Waisfeld, B.G.. 1999. Acritarch biostratigraphy of the
upper Tremadoc - Arenig of the Eastern Cordillera, northwestern Argentina:
relationships with graptolite and trilobite faunas. Bollettino
della Societa Paleontologica Italiana. 38 (2-3): 267-286.
Rubinstein,
C.V. and Vaccari, N.E. 2001. Palynomorphs of the Ordovician-Silurian boundary in
the Salar del Rincón Formation, Argentine Puna. First Meeting of the
C.I.M.P. Spores and Pollen Subcommission, National University
of
Ireland, Cork, Ireland. Abstracts, pp. 40-41.
Servais,
T. and Fatka, O. 1997. Recognition of the Trans-European Suture Zone (TESZ) by
the palaeobiogeographical distribution pattern of early to middle Ordovician
acritarchs. Geolical Magazine 134 (5): 617-625.
Servais,
T. and Molineux, S. 1997. The Messaoudensis-Trifidum acritarch
assemblage (Ordovician: late Tremadoc-early Arenig) from the subsurface of Rugen
(Baltic Sea, NE Germany). Palaeontographia Italica. 84:
113-161.
Staplin,
F.L. 1969. Sedimentaryorganic
matter, organic metamorphism, and oil and gas occurrence. Bulletin of
Canadian Petroleum Geology. 17: 47-66.
Steemans,
P., Higgs, K.T. and Wellman, P. 2000. Cryptospores and trilete spores from the
Llandovery, Nuayyim-2 borehole, Saudi Arabia. Al-Hajri, S & Owens B.
(eds.). GeoArabia Special Publication 1. Gulf PetroLink, Bahrain. pp.
92-115.
Tongiorgi,
M., Di Milia, A., Le Fort, P. and Gaetani, M. 1994. Palynological dating
(Arenig) of the sedimentary sequence overlying the Ishkarwaz Granite (upper
Yarkhun valley, Chitral, Pakistan). Terra Nova 6: 595-607.
Toro,
B.A. 1996. Implicancias paleogeográficas del hallazgo de Baltograptus
turgidus (Lee) y B. Kunmingensis (Ni) (Graptolithina) en el Arenigiano
Temprano del Noroeste de Argentina. XIII Congreso Geológico Argentino y III
Congreso de Exploración de Hidrocarburos. 5: 27-38.
Turner,
J. C. 1964. Descripción
geológica de la hoja 15c, Vinchina. Boletín del Instituto Nacional de
Geología y Minería. 100: 93.
Vavrdova,
M. 1974. Geographical differentiation of Ordovician acritarch assemblages in
Europe. Review of Paleobotany and Palynology. 18: 171-175.
Vavrdova,
M. 1997. Early Ordovician provincialism in acritarch distribution. Review of
Palaeobotany and Palynology. 98: 33-40.
Vecoli,
M. 2000. Palaenvironmental interpretation of microphytoplankton diversity trends
in the Cambrian- Ordovician of the northen Sahara Platform. Palaeogeography,
Paleoclimatology, Palaeocology. 160:329-346.
Volkheimer,
W. 1978. Faunas de quitinozoos del Paleozoico Inferior de la Argentina. Reunión
Nacional del Paleozoico de Argentina. Universidad Nacional de Tucumán.
15-18.
Volkheimer,
W., Cuerda, A.J. and Melendi, D.L. 1980a. Quitinozoos de la Formación
Gualcamayo en su localidad tipo al sudoeste de Guandacol, Precordillera de La
Rioja, República Argentina. II Congreso Argentino de Paleontología y
Bioestratigrafía y I Congreso Latinoamericano de Paleontología. Actas. 1:
23-36.
Volkheimer,
W., Melendi, D.L. and Aceñolaza, F.G. 1980b. Una microflora ordovícica de la
Formación Mojotoro, provincia de Salta. Revista de la Asociación Geológica
Argentina. 35
(3): 401-416.
Waisfeld,
B.G. 1993. Concentraciones
fosilíferas autóctonas y parautóctonas en la Fm. Acoite (Ordovícico
temprano), Cordillera Oriental, Provincia de Jujuy. Consideraciones tafonómicas
y paleoecológicas. XII Congreso Geológico Argentino y II Congreso de
Exploración de Hidrocarburos. 2: 296-303.
Waisfeld,
B.G. 1996. Revisión
de la Zona de Hoekaspis schlagintweiti Harrington y Leanza, Ordovícico
del noroeste de Argentina. 12º Congreso Geológico de Bolivia. 3:
915-921.
Wang,
Y., Li, J. and Wang, I. 1997. Latest Ordovician cryptospores from southern
Xinjiang, China. Review of Palaeobotany and Palynology. 99: 61-74.
Wellman,
C.H. 1999. Ordovician land plants: evidence and interpretation. Acta
Universitatis Carolinae. (Geologica).
43 (1-2): 275-277.
Recibido:
15 de agosto 2002
Aprobado: 18 de diciembre de 2002